СЕЛЬСКОХОЗЯЙСТВЕННАЯ БИОЛОГИЯ, 2008, № 1, с. 96-111
В порядке обсуждения
Филогенез и онтогенез растений
УДК 633.491:581.1:581.4:575.85/.89
ФИЛОГОГЕНЕТИЧЕСКАЯ МОДЕЛЬ РАЗВИТИЯ ЖИЗНЕННОЙ
ФОРМЫ ЦВЕТКОВОГО РАСТЕНИЯ И ТЕОРИЯ СЕЛЕКЦИОННОГО ПРОЦЕССА
(на примере картофеля)
Н.А. ЖУРАВЛЕВА
На основании данных литературы и собственных исследований обсуждается возможность применения эволюционного подхода к созданию теории сорта и моделей высокопродуктивных сортов картофеля. Предпринята попытка оценить процесс клубнеобразования, как результат филогенетического развития жизненных форм цветковых травянистых растений, структурной единицей которых является побег. Появление клубня в его составе рассматривается как заключительный этап эволюции побега травянистых двудольных растений, исходной видовой формой которых был кронообразующий побег, завершающей — побег с интенсивным клубнеобразованием. Предложена модель этого процесса, которая позволяет объяснить происхождение основных селекционных признаков картофеля.
Ключевые слова: филогенез, жизненная форма, травянистый побег, клубнеобразование, дикие виды картофеля, сорта Solanumtuberosnm L.
Key words: phylogenesis, growth form, herbaceous shoot, tuberization, wild potato species, Solanum tuberosnm L. varieties.
Повышение потенциала продуктивности сельскохозяйственных растений относится к основным задачам селекции. Однако создание сорта, как отмечал Н.И. Вавилов (1), есть не что иное, как «эволюция, направляемая волей человека». Поэтому совершенствование методологии и технологии селекции, в том числе приемов генетической инженерии, связано как с расширением возможностей применения молекулярно-генетических методов и ДНК-технологий (2), так и с разработкой ряда тех основных вопросов теоретической биологии растений, которые определяют современное состояние теории эволюции растений как основы теории сорта. К таким вопросам эволюционной морфологии относится взаимодействие генотипа, контролирующего тот или иной признак, и среды, и в частности процессы образования биомассы и формообразования побега как жизненной формы цветковых травянистых растений (3-7).
Ключевой проблемой остается механизм эволюции генома, начавшийся с возникновения 3,5 млрд лет назад фототрофии (8) и приведший к последовательному появлению ряда жизненных форм растений: Thallobionta (Талломные, одно- и многоклеточные водоросли)®Procormobionta (Предпобеговые моховидные)®Cormobionta archegoniatae (Побеговые архегониальные, в эволюционной последовательности плауны, хвощи, папоротники, голосеменные)®Cormobionta gynoeciatae (Побеговые пестичные, или цветковые) (9). То есть после выхода 400-450 млн лет назад на сушу растения прошли все стадии изменения жизненной формы: от слабодифференцированного таллома, состоящего из листо- и корнеподобных филлоида и ризоида (ранние моховидные), до настоящего побега, точнее, корнепобега (10), у которого каждый орган (корень, стебель, лист) максимально специализирован (архегониальные) и единственным донором пластических веществ стали фотосинтезирующие клетки ассимиляционно-транс-пирационного органа — листа. Последний крупный этап развития жизненной формы растений, начавшийся примерно 140-144 млн лет назад, ознаменовался появлением цветковых, корнепобег которых описывается формулой: корень-стебель-лист-цветок/плод. У цветковых возникли многочисленные иерархии генетически закрытых популяций разного объема — семейства и роды, а в пределах последних — виды, появление которых представляет, по словам Ч. Дарвина (11), «ужасающую тайну», которая не раскрыта и сегодня. Улучшенные разновидности ничтожно малой их доли и составляют сортовой фонд человечества.
Эволюция корнепобега цветковых, особенно последние 25 млн лет, протекала очень быстро. Отметим, что именно на этот период (миоцен), составивший 0,7 % времени существования фототрофии, пришлись крупнейшие геологические события, сформировавшие поверхность Земли в ее современном виде (передвижение континентов, поднятие многих горных систем, изменявших направление океанических и воздушных потоков), что обусловило геологически быстрые смены климата. Произошла его аридизация и похолодание, установились сезонные изменения, резко снизилось количество CO2 в воздухе (12) (на некоторых территориях содержание воды и теплообеспеченность в почве достигали лимитирующих значений). Для растений это означало, что на разных участках суши изменялась ресурсообеспеченность жизнедеятельности, что приводило к изменению количества и качества производимых листом ассимилятов и вторичных метаболитов, которые использовались для создания наиболее экономичных видовых ризопобеговых форм. В результате в ряду сменяющихся форм, которые в разные геологические эпохи на разных территориях получали преимущественное развитие (деревья, кустарники, полукустарники, однолетние и многолетние травы) (13), биомасса особи постепенно уменьшалась. У финальной формы корнепобега (многолетней травянистой)образовались различные оптимизированные варианты: от максимально редуцированного с формулой корень-стебель-цветок/плод, где функцию листа начинал исполнять стебель (появление видов сем. Coctaceae в безводных и жарких местообитаниях) до так называемого «сверхполного» — (корень-стебель-лист-цветок/плод)-(столон-клубень), который возник в условиях короткого дня и сниженных водо- и теплообеспеченности и пример которого — появившиеся на Панамском перешейке и в Андах (4000 м над уровнем моря) в роде Solanum (сем. Solanaceae) более 170 видов клубнеобразующего паслена (картофеля), объединенных по этому признаку в секцию Tuberarium (14).
Вопросы молекулярной и эволюционной геномики растений (15), их физиологии и теории продукционного процесса (16-18), а также биоморфологии растений (19), связанные с расшифровкой механизма филогенеза корнепобега цветковых растений как интегрального явления, в основе которого лежит развивающийся геном, пока далеки от решения. Можно предположить, что произошедшие изменения в растительном геноме, затрагивающие деятельность фотосинтезирующих клеток листа, были адекватны изменению ресурсообеспечения корнепобега в целом, а также той доле ресурсов, которая поступала в ассимиляционную паренхиму листа. В результате изменялись масса, объем и форма этих клеток, а затем и все формируемые с использованием образованных ими пластических веществ структуры растительного организма. Однако механизм генетической регуляции морфогенеза и передачи информации по цепочке интегративных уровней клеточной иерархии у побеговых цветковых растений, который обеспечивает развитие конкретного комплекса морфологических признаков на уровне вида (а в его рамках — признаков сорта), пока также неясен (20).
Все основные виды сельскохозяйственных растений относятся к цветковым, а наиболее важные из них (пшеница, кукуруза, овощные культуры, картофель и др.) имеют форму травянистого корнепобега. Дальнейшее изменение формы корнепобега за счет увеличения массы одного из его органов и составляет суть работы селекционера при создании высокопродуктивного сорта. В таком контексте корнепобег травянистых цветковых растений можно представить как (корень)-(стебель-лист-цветок/плод)-(столон-клубень), причем баланс этих функционально различающихся и определенным образом структурированных частных биомасс устанавливался в результате филогенетического развития в условиях изменявшегося ресурсообеспечения в месте обитания. Корнепобег с преимущественным развитием какого-то одного целевого органа соответствует определенной хозяйственной группе — корнеплоды, листовые, стеблевые и ягодные овощи, зерновые, картофель. Естественно сложившиеся в филогенезе балансы биомасс побега характеризуют так называемые дикие формы этих видов (21). В результате прямого отбора или отбора гибридов появились формы корнепобегов с гипертрофией соответствующего целевого органа, то есть их высокопродуктивные разновидности — сорта. Алгоритм создания корнепобега, целевой функцией которого является максимальная масса того или иного органа (22), может полностью или частично совпадать с историческими тенденциями его развития или быть прямо противоположным, поскольку все полезные с точки зрения человека признаки были отобраны эволюцией и формировались на протяжении многих миллионов лет как целостная общая корреляционная система, вследствие чего насильственное изменение одного из них может повлечь за собой каскадное изменение других, причем необязательно в нужную для селекционера сторону (23).
Теоретические модели, реконструирующие механизмы последовательной эволюции жизненных форм фотоавтотрофов вплоть до появления формы травянистого корнепобега, позволят оценивать возможности дальнейшего увеличения биомассы тех органов, которые в селекции являются целевыми: плодов (виды сем. Gramineae, Cucurbita-ceae), листьев, стебля или цветка (разные виды из сем. Cruciferae: Brassica capitata, B. gongyloides, B. botrytis); корня (виды сем. Chenopodiaceae) или клубней, как у вида Solanum tuberosum. При этом эффективное создание сортовых корнепобеговых форм того или иного вида представляет собой своего рода «сверхэтап» эволюционного развития ризопобега, характерного только для цветковых травянистых растений, который связан с волевым изменением его естественно образовавшейся формы в полезную для человека сторону. Теория сорта и селекционного процесса должна не только раскрывать факторы, инициировавшие появление базовых морфологических признаков на уровне вида в филогенезе, но и давать полное представление о физико-химическом механизме прохождения этой информации от целевого гена через клетку, ткань, орган до целостного многоклеточного организма. Такие сведения необходимы при планировании селекции высокопродуктивного или скороспелого районированного сорта любого вида сельскохозяйственных растений, адекватного условиям тепло-, свето- и водообеспечения региона-заказчика как основным нерегулируемым факторам среды, особенно если такой сорт предполагается получить путем прямого изменения генома. Однако создание моделей филогенеза корнепобега травянистого цветкового растения (140 млн лет) либо, что еще лучше, моделей, охватывающих более крупные периоды развития наземных растений (440 млн лет) или фотосинтезирующей массы вообще (3,5 млрд лет), которые смогли бы стать основой для создания различных сортовых моделей у видов цветковых травянистых растений, остается одним из основных и пока что не решенных, а главное, мало обсуждаемых в специальной литературе вопросов современной теоретической биологии.
К а р т о ф е л ь к а к о б ъ е к т д л я с о з д а н и я м о д е л и в ы с о к о п р о д у к т и в н о г о и с к о р о с п е л о г о с о р т а. Возникновение в фило- или онтогенезе побега любого элемента всегда связано либо с генетически детерминированным появлением и постепенным увеличением объема особым образом структурированной биомассы, либо с перераспределением биомассы или формообразованием соответствующих более частных структур в составе уже имеющейся. В структуре побега у растений картофеля в отличие от видов с более простой формой, не образующих клубни, имеется столонно-клубневый элемент. Цель селекции картофеля — увеличение только массы клубней, механизм образования которых (за исключением отдельных деталей) пока неясен (24-26).
Механизм клубнеообразования обычно изучают на примере S. tube-rosum или других интенсивно образующих клубни видов — Helianthus tuberosus, Oxalis tuberosa и т.д. и только как онтогенетический этап их развития (24). Подобный подход оставляет ряд вопросов, решение которых крайне необходимо для задач селекции. Например, как изменится потенциал клубнеобразования S. tuberosum при использовании в селекционном процессе других видов картофеля; чем обусловлено структурирование биомассы клубней, то есть их размер и форма, выравненность, кучность расположения вокруг основания побега; как связана фотосинтетическая активность листьев как источника пластических веществ для формирования побега в целом и появление на определенном этапе фило- и онтогенеза сначала столонов, а затем клубней. Из-за неразрешенности этих вопросов при создании сорта картофеля, его апробации или промышленном использовании, когда известны условия выращивания и возможна оценка любого морфофункционального признака стебле-листовой части побега, характеристика клубневой массы часто оказывается непредсказуемой (27). Если появление столонно-клубневого элемента с преимущественным развитием клубней рассматривать как заключительный этап филогенетического развития травянистого побега цветковых растений (Cormobionta gynoeciatae) и иерархически еще более высоких таксонов наземных растений — класса Cormobionta и царства Plantae, то представляет интерес определение общего принципа перехода одной формы побега в другую вплоть до столонно-клубневой (появление в роде Solanum секции Tubera-rium)и ее превращения у некоторых видов этой секции в интенсивно клубнеообразующую (S. andigenum, S. tuberosum и др.).
М о р ф о г е н е з п о б е г а ц в е т к о в о г о т р а в я н и с т о г о р а с т е н и я к а к с л е д с т в и е и з м е н е н и я б а л а н- с а б и о м а с с ы е г о э л е м е н т о в (описание модели). В основе построения модели лежит паритет ботанических и физиологических концепций развития побега: представление о размещении относительно поверхности земли боковых почек побега как одного из условий сохранения вида при ухудшении водо- и теплообеспеченности (система Раункиера) (28) и представление о донорно-акцепторных отношениях при распределении ассимилятов в системе побега (29). При построении модели в качестве исходной структурной единицы был принят побег, включающий элементы согласно формуле: стебель-лист-цветок/плод. Все возникающие на нем типы боковых побегов расценивались, во-первых, как дополнительные элементы, создающие вместе с основным побегом его более сложные видовые формы, и, во-вторых, как производители дополнительных ассимилятов, которые идут на увеличение биомассы побега и массы семян, обеспечивающих возможность генеративного возобновления вида. Видовые формы побега ранжировали по высоте закладки на стебле главного побега (от h до –h) оснований боковых побегов (рис. 1), образующих функциональный ряд, в котором боковые побеги постепенно утрачивают первоначальную фотосинтезирующе-репродуктивную роль: от повторяющих схему основного (стебель-лист-цветок/плод) через фотосинтезирующие вегетативные, развивающиеся по схеме (стебель-лист), до не производящих, а только потребляющих пластические вещества и состоящих из одного элемента (стебель), то есть подземного столона с разной степенью утолщения на конце. При сопоставлении этих форм можно обнаружить сходство с динамическими рядами некогда дивергировавших травянистых побеговых форм двудольных и филогенетически более молодой таксономической группы однодольных растений (30) в соответствии с законом гомологических рядов в наследственной изменчивости (31). Их исходной формой, образовавшейся при оптимальной водо- и теплообеспеченности, был кронообразующий побег с характерной многопорядковостью осей, длинными междоузлиями и крупной листовой пластинкой (см. рис. 1, а, б). Соответствующие боковые побеги создают дополнительные ассимиляты, обеспечивая формирование мощной долгоживущей стебле-листовой части побега и массы семян, достаточной для воспроизведения вида только генеративным путем.
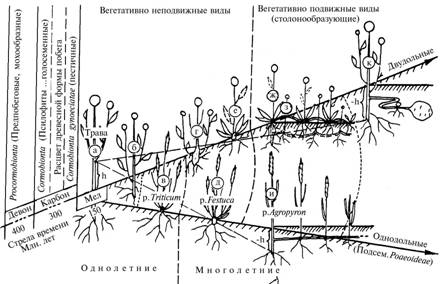
|
Рис. 1. Схема развития побега травянистых покрытосеменных растений по изменению высоты закладки на основном стебле (от h до -h) оснований боковых побегов: а (б) — исходная кронообразующая форма; в — появление зоны кущения; г — появление многолетнего корневища; д, е — розеточная и полурозеточная формы побега, ж-и — появление надземного и подземного столонов и образование на них дочерних ризопобегов; к — образование подземного столона и клубня (реконструкция). |
Аридизация климата, протекавшая на фоне высоких температур, способствовала появлению слабооблиственного побега меньшего размера и снижению массы листьев, вследствие чего не обеспечивалось нужное развитие репродуктивного элемента. Появление в основании стебля такого побега так называемой зоны кущения, в которой часть боковых побегов иногда становилась генеративной (см. рис. 1, в), компенсировало этот процесс. При расширении ареала таких видов в сторону территорий со сменой сезонов года и усилением континентальности климата (32) произошло постепенное перемещение зоны кущения побега в при- и подземную часть стебля. На подземной части стали располагаться почки ежегодного возобновления побега, что и определило появление в подобном климате у цветковых травянистых растений многолетней формы побега, у которой надземная стебле-листовая часть, ежегодно возобновляясь, за один вегетационный сезон полностью завершала жизненный цикл, а подземная стеблевая за счет формирования корневища становилась долгоживущей (десятки и даже более сотни лет) (см. рис. 1, г-ж). При этом изменялась и так называемая подвижность побегов (28). На начальных этапах эволюции побега у цветковых травянистых растений, протекавших при оптимальных (двудольные), а затем близких к оптимальным условиях водо- и теплообеспеченности (однодольные), травянистые виды существовали только в форме вегетативно неподвижных побегов (см. рис. 1, б-е) и размножались только семенами, что обеспечивало им сохранение и процветание во влажных и теплых (виды овощных культур) или относительно сухих и теплых (хлебные злаки) местообитаниях, ставших соответствующими центрами видообразования (21).
Дальнейшее развитие многолетних травянистых форм побега в жестких условиях резко континентального климата привело к уменьшению надземной массы побега в целом, главным образом за счет укорочения междоузлий, то есть уменьшения массы стебля (33). Этот процесс определил последовательное появление сначала полурозеточной, а затем и розеточной формы побега (см. рис. 1, г-е), у которой баланс биомассы элементов оказался максимально смещен в сторону ассимилирующей листовой части.
С появлением розеточной формы дальнейшая трансформация побега посредством простого перераспределения биомасс в системе стебель«лист«цветок/плод оказалась исчерпанной, поэтому следующий этап развития цветковых травянистых растений был связан с усложнением формы самого побега за счет появления принципиально нового элемента — столона, то есть бокового плагиотропного стебля с располагающимися на нем почками, из которых развивались дочерние корнепобеги (см. рис. 1, ж-к). Возникновение столона у некоторых видов растений в неблагоприятных условиях среды стало мощной альтернативой семенного возобновления побега, поскольку имело место своего рода естественное клонирование. При этом произошло не только ускорение роста популяций этих видов благодаря образованию многочисленных генокопий материнских побегов, возникших посредством семенного размножения, но и быстрое продвижение таких вегетативных «клонов» к границам ареала вида, которые определялись сочетанием основных лимитирующих факторов — водо-, свето- и теплообеспеченности.
Однако и в этом ряду можно наблюдать постепенный переход столонов из надземной части побега в подземную. На начальных этапах эволюции столона как у дву-, так и у однодольных растений он еще участвовал в создании дополнительных ассимилятов благодаря образованию дочерних побегов, повторяющих схему основного побега: стебель-лист-цветок/плод (см. рис. 1, ж-и). Но у двудольных такой столон, пройдя все стадии развития (см. рис. 1, ж и з), постепенно превращался в не имеющий выхода на поверхность полностью потребляющий элемент — подземный стебель (см. рис. 1, к). Его концевая часть постепенно обретала форму клубня, на котором стал располагаться основной запас почек для вегетативного возобновления. При этом собственно столон способствовал перемещению этих почек на то или иное расстояние от основания побега. Надземная же часть снова обрела типичную кронообразующую форму, которая, однако, стала более короткоживущей в соответствии с новыми условиями теплообеспеченности и продолжительностью сезона вегетации в местообитании вида.
Во многих семействах двудольных возникновение клубнеобразующих видов было единичным явлением. Появление же в роде Solanum примерно 170 видов клубнеобразующего паслена, или картофеля, — событие уникальное, поскольку в этом случае видообразование протекало только в пределах отличной от остальных видов этого рода столоно-клубнеобразующей формы побега, единой формулы цветка и только по типу увеличения плоидности (2n — 24, 36, 48, 60 и 72) (27). Общая длина многочисленных столонов у некоторых видов картофеля (например, S. berthaultii) может достигать 20 м и более, то есть даже одна возникшая полиплоидная особь как родоначальница нового вида картофеля вследствие особенностей жизненной формы и образования клонов быстро заселяла соответствующий ареал. На территории Центральной и Южной Америки (центры видообразования картофеля) специфическим фактором, обусловливающим появление/непоявление в подземной части побега столонов и клубней, была длина дня (световой ресурс), которая играла существенную роль в становлении жизненных форм у видов картофеля (26) и корректировке их ареалов, определяемых основными лимитирующими факторами эволюции растений — водо- и теплообеспеченностью.
Таким образом, появление у некоторых видов травянистых двудольных растений столонно-клубневого элемента стало одним из замечательных эволюционных приобретений, способствовавшим увеличению численности их популяций и более быстрому освоению ареала. Однако при этом возникла необходимость содержать в составе побега дополнительное количество акцепторной подземной биомассы, которая у отдельных видов картофеля (например, S. commersonii) может составлять до 30-40 %. Процесс завершился возникновением столонно-клубневой формы побега у видов разных семейств, а также появлением в роде Solanum секции Tuberarium. В пределах указанной секции продолжалось развитие такой формы побега посредством перераспределения биомассы столона и клубней в пользу последних. Начальному этапу этого процесса соответствует побег с преимущественным развитием столонов, характерный для диких видов картофеля, замыкает динамический ряд побег растений вида S. tuberosum с активным клубнеобразованием.
И з м е н е н и е б а л а н с а б и о м а с с ы о р г а н о в р а с т е н и я к а к о т р а ж е н и е т е н д е н ц и й р а з в и т и я у в и д о в к а р т о ф е л я. Каков механизм появления в составе травянистого побега у видов двудольных растений подземного столона, а затем и клубня? Какое количество акцепторной биомассы побега может обеспечивать его надземная ассимилирующая часть? До какой степени у видов картофеля подземная акцепторная биомасса может дифференцироваться на столоны и клубни? Поставив перед собой эти вопросы, мы изучили накопление биомассы элементами побега у форм картофеля с разной способностью образовывать клубни — семи диких видов из коллекции Всероссийского НИИ растениеводства им. Н.И. Вавилова и восьми разновидностей (сортов) вида S. tuberosum. Растения выращивали из черенков в однопобеговой форме в индивидуальных контейнерах в теплице; повторность опыта 10- (сорта S. tuberosum) и 3-5-кратная (дикие виды). В конце августа в период
полного формирования побега определяли сухую массу каждого элемента у каждой особи (ошибка среднего 5-8 %). Ряд форм побега ранжировали по филогенетическому возрасту (дикие виды®сорта S. tuberosum) и в каждой из этих групп — по степени развития клубня.
1. Структура побега и его подземной части у растений диких видов Solanum и сортов S. tuberosum
|
Показатель |
Дикий вид Solanum(n= 3-5) |
Сорт S. tuberosum (n = 10) |
S. spegazzini |
S. demissum |
S. chacoense |
S. pinnatisectum |
S. andigenum |
S. berthaultii |
S. commersonii |
x1 |
Колпашевский |
Волжанин |
И. Барская |
Полет |
Тулунский |
Адретта |
Сосновский |
Приекульский |
х2 |
x1/x2 |
С т р у к т у р а п о б е г а |
Масса побега, г: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
надземная часть (Н) |
237 |
324 |
454 |
366 |
345 |
1047 |
179 |
424 |
1030 |
842 |
823 |
554 |
822 |
449 |
804 |
682 |
738 |
1,7 |
подземная часть (П) |
19 |
50 |
197 |
106 |
103 |
307 |
93 |
125 |
478 |
413 |
723 |
585 |
530 |
465 |
627 |
610 |
554 |
4,4 |
общая (О) |
256 |
374 |
651 |
472 |
448 |
354 |
290 |
549 |
1508 |
1255 |
1546 |
1139 |
1352 |
914 |
1431 |
1192 |
1292 |
2,3 |
Н/П |
12,5 |
6,4 |
2,3 |
3,4 |
3,3 |
3,4 |
2,1 |
4,8 |
2,1 |
2,0 |
1,1 |
0,9 |
1,5 |
0,9 |
1,3 |
0,9 |
1,3 |
— |
О/П |
13,5 |
7,5 |
3,3 |
4,4 |
4,4 |
3,7 |
3,1 |
5,7 |
3,1 |
3,0 |
2,1 |
1,9 |
2,5 |
2,0 |
2,3 |
1,9 |
2,3 |
— |
Доля в массе побега, %: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
подземная часть |
92 |
87 |
70 |
78 |
77 |
77 |
68 |
78 |
68 |
67 |
53 |
49 |
61 |
49 |
56 |
49 |
56 |
|
надземная часть |
8 |
13 |
30 |
22 |
23 |
23 |
32 |
22 |
32 |
33 |
47 |
51 |
39 |
51 |
44 |
51 |
44 |
|
С т р у к т у р а п о д з е м н о й ч а с т и п о б е г а |
Масса, г: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
столоны (С) |
19,4 |
50,2 |
197,0 |
106,0 |
90,0 |
217 |
17,5 |
99,6 |
11,9 |
7,3 |
7,2 |
4,7 |
4,5 |
3,2 |
4,3 |
3,0 |
5,8 |
|
клубни (К) |
0 |
0 |
0 |
0 |
13,6 |
90,2 |
75,5 |
25,6 |
446 |
406 |
716 |
480 |
528 |
462 |
623 |
607 |
533 |
|
всего |
19,4 |
50,2 |
197,0 |
106,0 |
103,6 |
307,2 |
93,0 |
125,2 |
458 |
413 |
723 |
485 |
532 |
465 |
627 |
610 |
539 |
|
Н/К |
¥ |
— |
— |
— |
24,5 |
11,6 |
2,6 |
13,2* |
2,3 |
2,1 |
1,2 |
1,1 |
1,6 |
1,0 |
1,3 |
1,0 |
1,4 |
|
С/К |
¥ |
— |
— |
— |
6,60 |
2,40 |
0,22 |
3,10* |
0,026 |
0,018 |
0,010 |
0,010 |
0,008 |
0,07 |
0,07 |
0,005 |
0,012 |
|
Доля в массе побега, %: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
столоны |
100 |
100 |
100 |
100 |
87 |
71 |
19 |
82 |
2,6 |
1,8 |
1,0 |
1,0 |
0,7 |
0,7 |
0,5 |
1,1 |
1,1 |
|
клубни |
0 |
0 |
0 |
0 |
13 |
29 |
81 |
18 |
97,4 |
98,2 |
99,0 |
99,0 |
99,3 |
99,3 |
99,5 |
98,9 |
98,5 |
|
П р и м е ч а н и е. Прочерк означает, что показатель не определяли, пропуск — отсутствие показателя; x1 и x2 — среднее соответственно для видов Solanum и сортов S. tuberosum.
* Среднее по показателям у трех видов, сформировавших клубни. |
Средняя масса побега у сортов S. tuberosum (1292 г), масса надземной (738 г) и подземной его части в целом (554 г) по сравнению с таковыми у растений диких видов (соответственно 549, 424 и 125 г) возрастала соответственно в 2,3; 1,7 и 4,4 раза. При этом отмечалось относительное уменьшение доли надземной части (с 78 до 56 % соответственно у диких видов по сравнению с сортами S. tuberosum) и увеличение — подземной (соответственно с 22 до 44 %) (табл. 1).
Увеличение доли биомассы подземной части побега в филогенезе форм картофеля сопровождалось перераспределением биомассы между столонами и клубнями в пользу клубней: у растений диких видов на долю столонов приходилось 82, клубней — 18 %, у сортов S. tuberosum — соответственно 1,1 и 98,9 %; у некоторых диких видов картофеля (S. spegazzini, S. demissum) это соотношение достигало соответственно 100 и 0 %, а у сорта Приекульский (S. tuberosum) — соответственно 0,5 и 99,5 % (см. табл. 1). То есть в пределах секции Tuberarium отмечается чрезвычайно высокая изменчивость этого признака: от принципиальной возможности формировать только столоны (некоторые дикие виды картофеля) до образования подземного побега, состоящего преимущественно из клубней (S. tuberosum).
Рассмотрим, как в филогенезе побега у видов картофеля изменялось соотношение биомассы его донорных (надземных ассимилирующих) и потребляющих пластические вещества подземных органов, а также эффективность первых по обеспечению формирования и поддержанию вторых. Для этого введем понятия основного структурообразующего (соотношение биомассы надземной и подземной частей побега, характеризующие его форму на уровне секции Tuberarium) и уточняющего (соотношение биомассы столонов и клубней, характеризующее форму побега на уровне вида) балансов (табл. 2) и сопоставим полученные оценки с характеристикой структуры побега (см. табл. 1). Как видно, увеличение общей массы побега у сортов S. tuberosum (в среднем в 2,3 раза) происходило в основном за счет подземной части, у которой масса по сравнению с аналогичным показателем для диких видов картофеля возросла в 4,4 раза, тогда как у надземной — лишь в 1,7 раза. Такое значительное увеличение биомассы подземной части побега у эволюционно более молодого вида стало результатом двух противоположных тенденций — уменьшения массы столонов в 17,2 раза и увеличения массы клубней в 20,8 раза (см. табл. 2). При этом усиление столонообразования у представителей диких видов (оно в 17,2 раза выше по сравнению с S. tuberosum), в свою очередь, коррелировало с крайне низкой степенью клубнеобразования, а в ряде случаев даже с его отсутствием (см. табл. 1).
2. Изменение структуры побега у видов картофеля в филогенезе
|
Вид Solanum |
Побег |
Часть побега |
надземная |
подземная |
столоны |
клубни |
С р е д н я я м а с с а, г |
Дикие виды (х) |
549 |
424 |
125 |
99,6 |
25,6 |
S. tuberosum (у) |
1292 |
738 |
554 |
4,8 |
533,8 |
С о о т н о ш е н и е м а с с ы п о б е г а и е г о ч а с т е й |
x/у |
– |
– |
– |
17,2 |
– |
у/х |
2,3 |
1,7 |
4,4 |
– |
20,8 |
П р и м е ч а н и е. Прочерк означает величину показателя < 1. |
При менее значительном приросте биомассы надземной части побега по сравнению с подземной в филогенезе форм картофеля повышалась эффективность участия первой в образовании второй, надо полагать, вследствие соответствующего изменения гормонального баланса. Так, у более ранних форм (дикие виды) в создании 1 г массы подземной части побега в среднем участвует 4,8, а у поздней (S. tuberosum) — 1,3 г надземной (см. табл. 1). В отношении формирования клубней тенденция становится еще более заметной — соответственно 13,2 и 1,4 г (см. табл. 1).
Подобные изменения в структуре побега и эффективности работы его надземной части позволяют сделать вывод, что в филогенезе все большая доля постепенно увеличивающейся подземной биомассы как бы «перетекала» в клубень. Более того, чрезвычайно низкое соотношение массы столонов и клубней у наиболее молодого вида S. tuberosum (0,012) по сравнению с дикими видами (3,10) и усиление этой тенденции в ряду сортов S. tuberosum (от 0,026 до 0,005) (см. табл. 1) свидетельствуют, что у этого вида при сложившейся степени развития стебле-листовой массы возможность дальнейшего увеличения массы клубней за счет столонов практически отсутствует.
Таким образом, структура побега, которая эволюционным путем естественно сформировалась в секции Tuberarium и далее видоизменилась в процессе селекции с обязательным участием S. tuberosum, как бы дважды себя исчерпала. То есть генетически детерминированный видовой предел образования биомассы надземной части побега создал барьер для увеличения общей подземной массы, что, в свою очередь, ограничило возможность увеличения биомассы клубней. Поэтому дальнейший рост последней возможен только за счет радикальных генетических преобразований, обусловливающих морфоанатомические и физиологические изменения стебле-листовой части побега у растений картофеля.
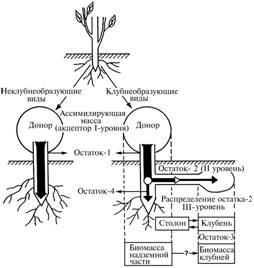
|
Рис. 2. Схема распределения ассимилятов в иерархии акцепторных элементов побега у видов, образующих и не образующих клубни. |
П о б е г к а к э в о л ю ц и о н и р у ю щ а я с и с т е м а с т р у к т у р н ы х э л е м е н т о в, с в я з а н н ы х д о н о р н о - а к ц е п
т о р н ы м и о т н о ш е н и я м и. Рассмотрим побег цветковых травянистых растений как поддерживаемую балансом прямых положительных и обратных отрицательных связей оптимизированную систему биомасс органов — доноров и акцепторов (34). В этой системе на определенном этапе филогенеза образуется дополнительный столонно-клубне-вый элемент как следствие появления сначала двух, а затем трех уровней распределения пула ассимилятов, синтезируемых в листьях (прямая положительная связь). Появление таких уровней определялось генетически детерминированным изменением соотношения морфогенных веществ (гормонов) в пуле ассимилятов (рис. 2).
Акцептором I уровня в такой системе являются листья, которые используют ассимиляты in situ. Предел увеличения массы листьев и всей надземной части побега определяется интенсивностью реакций фото- и биосинтеза (прямая положительная связь), а также теми структурными ограничениями, которые не позволяют этим элементам как акцепторам утилизировать все созданные ассимиляты (обратная отрицательная связь). К таким ограничениям относятся предельное число и объем ассимилирующих клеток, максимальная площадь листовых пластинок, диаметр стебля, число и длина междоузлий, детерминированные на уровне вида.
При образовании не востребованных надземной частью ассимилятов (остаток-1) они поступают в подземную часть стебля. Здесь возможно, во-первых, их продвижение в корень и даже выведение в почву (35). При этом часть ассимилятов расходуется на закладку в надземной части стебля боковых почек, из которых развиваются побеги следующих порядков, формируя разные варианты кронообразующего побега, однако в подземной части стебля столон не образуется (обозначим этот вариант формообразования побега как альтернативу «—»). Во-вторых, часть остатка-1 (остаток-2) может использоваться на заложение почек в подземной части стебля и формирование из них либо выходящих на поверхность производяще-потребляющих, либо подземных потребляющих боковых побегов. В этом случае крайний вариант развития в виде не выходящей на поверхность столонно-клубневой системы означает появление в составе побега акцепторной массы II уровня (альтернатива «+»).
Альтернативе «—» соответствует форма побега, характерная для большинства видов цветковых травянистых растений, а также не образующих клубни видов рода Solanum (S. melongena, S. muricatum, S. nigrum и др.). При альтернативе «+» (и только как крайний ее вариант) развивается побег, описываемый формулой: (лист-стебель-цветок/плод)-(столон-клубень). Такая форма побега встречается только у двудольных травянистых растений, к которым относятся примерно 12 % видов рода Solanum (сем. Solanaceae, секция Tuberarium) и единичные виды из других семейств — Helianthus tuberosus (сем. Compositae), Ipomea batatas (сем. Convolvulaceae), Oxalis tuberosa (сем. Oxalidaceae) и др.
Неполный учет функциональных и морфологических характеристик, определяющих величину и состав ассимилятов остатка-2, существенно затрудняет прогнозирование развития подземной части побега при оценке селекционного материала картофеля. Так, у S. commersonii и S. tuberosum (сорт Волжанин) при разной степени развития надземной части побега (соответственно 197 и 842 г) формируется ассимиляционный остаток-2, который в подземной части реализуется в равных долях от общего потока (соответственно 32 и 33 %). В то же время при одинаковом развитии надземной части побегов у S. berthaultii и S. tuberosum (сорт Колпашевский) (масса соответственно 1047 и 1030 г) предполагаемый остаток-2 у первого в 1,56 раза меньше, чем у второго (соответственно 307 и 478 г) (см. табл. 1).
Еще менее исследована взаимосвязь между развитием надземной части побега и собственно клубней как целевого признака в селекции картофеля (27). Согласно предлагаемой схеме, эта взаимосвязь формируется на III уровне распределения ассимилятов между аналогичными конкурирующими элементами — столоном и клубнем. При этом дистально расположенный клубень образуется за счет чрезвычайно подвижного остаточного количества ассимилятов — остатка-3. Окончательная схема распределения ассимилятов (и, соответственно, образования видовых форм побега у растений картофеля) следующая: (лист-стебель-цветок/плод)®(столон«клубень).
Кроме того, последовательность органогенеза (появление столонов®образование/отсутствие образования клубней) свидетельствует, что условием клубнеобразования является изменение баланса гормонов в составе остатка-2 при преимущественном накоплении цитокининов, абсцизовой кислоты и других ингибиторов роста (24-26) по сравнению с индукторами корне- и стеблеобразования — ауксинами и гиббереллинами. В этом случае процесс удлинения столона приостанавливается и за счет ассимиляционного остатка-3 начинается равномерное разрастание концевых паренхимных клеток, которое приводит к образованию клубня. Если индуцирующий клубнеобразование гормональный баланс в остатке-3 не формируется или формируется слишком поздно, происходит только удлинение столонов, и побег приобретает форму, характерную для более древних, так называемых диких видов картофеля.
Таким образом, с позиций донорно-акцепторной концепции в филогенезе побега единственным донором и первым акцептором ассимилятов является лист. Остальные элементы побега (стебель, цветок/плод, столон и клубень) формируются по остаточному принципу и как следствие изменений состава ассимилятов при разном ресурсообеспечении (вода, тепло, свет). Отметим, что клубнеобразующий побег развивался не только у части видов рода Solanum, но и в таксоне цветковых травянистых растений как таковых. На уровне генома описанные процессы были обусловлены разными генетическими событиями — увеличением объема и усложнением генома, изменением его структуры и функциональной активности (появление новых и дупликация имеющихся генов, обеспечивающих эволюционное преимущество, инактивация и переход генов, утративших эволюционное значение в сложившихся условиях среды, в категорию «молчащих») (36, 37). Накопление молчащих генов, в свою очередь, постепенно увеличивало видовой потенциал скрытой соматической изменчивости побега, которая в современных формах проявляется в виде атавизмов. Примером служит появление не свойственных побегу растений картофеля диатропно-ортотропных побегов―(сарментов), которые, как и столоны, развиваются только из подземных почек стебля, но выходят на поверхность, как у более древнего кронообразующего побега (38), или так называемых уродливых клубней у филогенетически молодого вида S. tuberosum вследствие восстановления у преобразующегося в клубень столона некогда утраченной способности к ветвлению, характерной для диких видов картофеля и клубнеобразующих видов из других семейств. Можно предположить, что последовательность экспрессии генов в органогенезе клубнеобразующего побега отражает порядок их появления в филогенезе. Такой подход феноменологичен, однако позволяет на единой основе рассматривать имеющиеся факты, выявлять ключевые точки проблемы клубнеобразования для экспериментальной проверки существующих гипотез и прогнозировать развитие хозяйственно ценных признаков побега, обусловливающих продуктивность и структуру клубневой массы.
М о д е л ь с о р т а к а р т о ф е л я к а к ч а с т н ы й с л у ч а й э в о л ю ц и о н н о й м о д е л и п о б е г а ц в е т к о в ы х т р а в я н и- с т ы х р а с т е н и й. Положения предложенной модели клубнеобразования могут быть использованы для создания теоретических моделей сортов сельскохозяйственных растений с заданными признаками (22), в том числе картофеля (39). В частности, это представление о побеге как развивающейся в фило- и онтогенезе системе элементов биомассы, в основе изменения которой лежит изменяющийся геном; о ведущей роли листовой массы; об аналогии столона и клубней и остаточном принципе их формирования. Клубень в такой системе рассматривается как «недоразвившийся» столон с характерными признаками стебля (способность к ветвлению, наличие верхушечной и боковых почек и междоузлий), а столон — как «удлинившийся» потенциальный клубень. Отсюда можно вывести ряд представлений о причинах формирования практически всех структурных признаков подземной и даже надземной части у растений картофеля — от формирования клубня («норма—не норма») до структуры куста при промышленном выращивании.
Так, форма клубней (один из ключевых признаков сорта картофеля) в рамках обсуждаемой модели будет выглядеть как способность или неспособность концевой части столонов к ветвлению. Действительно, ветвление столонов характерно для всех видов секции Tuberarium, но у S. tuberosum в отличие от некоторых более ранних видов картофеля (S. tarijense) и клубнеобразующих видов из других семейств (H. tuberosus) этот процесс в концевой части столона полностью подавлен, что и определяет правильную форму клубня у его сортов. Поэтому редкое появление у тех же сортов уродливых клубней можно рассматривать как неполное подавление признака в силу особенностей его генетического контроля. С этих позиций становится объяснимым и сортовое разнообразие формы нормального клубня у S. tuberosum в достаточно узких пределах изменчивости вида. Оно зависит от детерминированной на уровне сорта длины концевой части столона, составляющей предположительную зону клубня, клетки которой утрачивают способность к вытягиванию, в результате чего проксимальная часть столона развивается в обычном режиме, а концевой участок начинает увеличиваться в объеме. Чем он короче, тем сильнее выражен переход столона в клубень и совершеннее форма последнего (рис. 3).
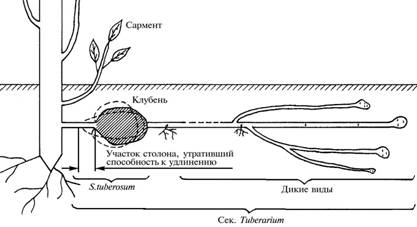
|
Рис. 3. Схема столоно- и клубнеобразования в секции Tuberarium. |
Согласно этой модели ряд важнейших селекционных признаков картофеля оказывается прямым следствием процессов, протекающих в листьях. Так, переориентация части потока ассимилятов с формирования надземной части (остаток-1) на формирование столонно-клубневого элемента (остаток-2) определяет срок заложения боковых почек в подземной части основного стебля, что обусловливает скороспелость сорта. Число таких почек, развившихся в столоны, и степень ветвления последних определяют число товарных (концевых) клубней и, следовательно, среднюю массу одного клубня, длина столонов — компактность расположения клубней вокруг основания стебля, топография и характер заложения почек возобновления побега на преобразующемся в клубень кончике столона (будущие так называемые «глазки») — расположение и глубину их заложения; время дозревания таких почек — лежкость клубней. Наконец, число почек возобновления на клубне и их состояние определяют число автономных побегов в кусте следующего года вегетации при промышленном выращивании картофеля.
При моделировании высокопродуктивного и скороспелого районированного сорта картофеля ключевыми являются два представления. Первое состоит в том, что общая масса побега определяется потенциалом его продуктивности, детерминированным на уровне вида (генотип), от которого прежде всего зависит величина и качество листовой массы, и наличием ресурсов жизнеобеспечения в условиях конкретного региона (среда), в которых этот генотип реализуется. Второе предполагает остаточный принцип формирования биомассы всех элементов побега (за исключением листьев), когда каждый онтогенетически более поздний элемент формируется на основе остаточного фонда ассимилятов, не востребованных ранее сформировавшимися.
На основании такого подхода при решении проблемы формирования клубней в связи с развитием надземной части побега в первую очередь встает вопрос о генетическом контроле образования ассимилятов остатка-1 (см. рис. 2). Оно, как отмечалось, определяется уникальным для каждого сорта сочетанием ассимилирующей способности клеток листа и анатомических характеристик надземной части побега, в том числе стебля, обусловливающих ее предельный депонирующий объем. Следующий вопрос — генетическая детерминация количества и состава ассимилятов остатка-2 (формирование столонно-клубневой части побега в целом) и лишь затем — максимизация накопления «целевого» ассимиляционного остатка-3, непосредственно идущего на образование клубней. Первый способ увеличения количества ассимилятов остатка-3 — создание побега с редуцированной надземной частью, у которого баланс биомасс элементов в надземной части максимально сдвинут в сторону листьев вследствие сближения междоузлий и усиления розеточности, а структурные ограничения депонирования ассимилятов листьями (предельный объем ассимилирующих клеток и площадь листовых пластинок) сложатся в наиболее поздние сроки. Это позволит получить максимальное количество остатка-1, но в течение более продолжительного времени, в связи с чем для реализации его избытка в качестве остатка-3 потребуются соответствующие ресурсы жизнеобеспечения (агротехника) и более длительный вегетационный период (базовый экологический фактор региона). Второй способ — максимально быстрое получение остатка-1 за счет более раннего ограничения развития депонирующего объема надземной части побега вследствие уменьшения размера клеток и листовой пластинки при сближенных междоузлиях.
Каждый из этих способов отражает алгоритм создания позднеспелого урожайного и раннеспелого, но менее урожайного сорта картофеля, причем в обоих случаях скороспелость сорта вступает в противоречие с урожайностью. Для преодоления отмеченного противоречия требуется создать форму побега с противоестественным сочетанием функциональных и структурных признаков: с одной стороны — максимальная активность фото- и биосинтетических процессов в клетках паренхимы, что является признаком естественно развивающегося мезоморфного листа и высокопродуктивного побега, с другой — максимально раннее развитие у элементов надземной части побега признаков ксероморфизма (мелкоклеточность, малая площадь листовой пластинки, сближенные междоузлия), которые обычно характеризуют низкопродуктивный побег (40-42). Потому при создании высокопродуктивного и одновременно раннеспелого сорта картофеля может быть осуществлен только компромиссный подход.
Таким образом, предложенная модель развития побега цветковых травянистых растений, отражая некоторые стороны эволюции продукционного процесса, может иметь и прикладное значение. Эволюционный подход открывает возможность более целенаправленного поиска условий проявления базовых признаков побега, в том числе появления и развития клубня, а также многих других селекционно значимых признаков у картофеля.
Автор выражает искреннюю благодарность Р.К. Саляеву, В.А. Давыдову, А.К. Глянько, И.А. Лукиной, В.К. Войникову, С.Г. Казановскому, и А.Т. Мокроносову за проявленный интерес и поддержку при написании статьи.
Л И Т Е Р А Т У Р А
- В а в и л о в Н.И. Селекция как наука. В сб.: Теоретические основы селекции. М., 1987: 7-68.
- Р о м а н о в Г.А. Генетическая инженерия растений и пути решения проблемы биобезопасности. Физиол. раст., 2000, 47(3): 343-353.
- У о д д и н г т о н К. Морфогенез и генетика. М., 1964.
- Ш н о л ь С.Э. Физико-химические факторы эволюции. М., 1979.
- С м и т Дж.М. Нерешенные проблемы эволюции. В сб.: Эволюция генома. М., 1986: 357-365.
- Ж у ч е н к о А.А. Адаптивный потенциал культурных растений. (Эколого-генетические основы). Кишинев, 1988.
- К у р б а т с к и й В.И. Цветковые растения ― новый этап в эволюции растительного мира. В сб.: Эволюционная биология. Томск, 2002: 165-78.
- К а л ь в и н М. Химическая эволюция. Молекулярная эволюция, ведущая к возникновению живых систем на Земле и на других планетах. М., 1971.
- Р е й в н П., Э в е р т Р., А й к х о р н С. Современная ботаника. М., 1990, т. 2.
- Н у х и м о в с к и й Е.Л. Основы биоморфологии семенных растений, М., 1997, т. 1.
- Д а р в и н Ч.Избранные письма. М., 1950.
- Б у д ы к о М.И. Эволюция биосферы. Л., 1984.
- Т а х т а д ж я н А.Л. Происхождение и расселение цветковых растений. Л., 1970.
- Б у к а с о в С.М. Систематика и география видов картофеля. В сб.: Генетика картофеля. М., 1973: 14-69.
- З е л е н и н А.В., Б а д а е в а Е.Д., М у р а в е н к о О.Е. Введение в геномику растений. Мол. биол., 2001, 35(5): 339-348.
- Н и ч и п о р о в и ч А.А. Фотосинтетическая деятельность растений как основа их продуктивности в биосфере и земледелии. В сб.: Фотосинтез и продукционный процесс. М., 1988: 5-28.
- М о к р о н о с о в А.Т. Физиология растений: проблемы развития и задачи ВОФР. М., 1988: 59-69.
- М о к р о н о с о в А.Т. Физиология растений на рубеже XXI века. Физиол. раст., 2000, 47(3): 341-342.
- Ю р ц е в Б.А. Жизненные формы: один из узловых объектов ботаники. В сб.: Проблемы экологической морфологии растений. М., 1976: 9-44.
- Х а в к и н Э.Е. Генетическая регуляция морфогенеза растений. Физиол. раст., 1998, 45(5): 463-777.
- В а в и л о в Н.И. Происхождение и география культурных растений. Л., 1987.
- И н г е - В е ч т о м о в С.Г. Генетика с основами селекции. М., 1989.
- Ж у р а в л е в а Н.А. Физиология травянистого сообщества. Принципы конкуренции. Новосибирск, 1994.
- У э р и н г Ф.Ф. Физиология клубнеобразования и роль фитогормонов. В сб.: Гормональная регуляция онтогенеза растений. М., 1984: 48-62.
- М о к р о н о с о в А.Т. Клубнеобразование и донорно-акцепторные связи у картофеля. В сб.: Регуляция роста и развития картофеля. М., 1990: 6-12.
- Ч а й л а х я н М.Х. Механизмы клубнеобразования у растений. В сб.: Регуляция роста и развития картофеля. М., 1990: 48-62.
- Б у д и н К.З. Генетические основы селекции картофеля. Л., 1986.
- С е р е б р я к о в а Т.И. Учение о жизненных формах растений на современном этапе. Итоги науки и техники (сер. «Ботаника»), 1972, 1: 84-169.
- М о к р о н о с о в А.Т. Фотосинтетическая функция и целостность растительного организма. М., 1983.
- В а к а р Б.А. Введение в филогению растительного мира. Минск, 1973.
- В а в и л о в Н.И. Закон гомологических рядов в наследственной изменчивости. Л., 1987.
- Ю с у ф о в А.Г. Лекции по эволюционной физиологии растений. М., 1985.
- Г о л у б е в В.Н. Эколого-биологические особенности травянистых растений и растительных сообществ лесостепи. М., 1965.
- Р о з е н Р. Принцип оптимальности в биологии. М., 1969.
- Л я л и н О.О., С о л о в ь е в Е.В. Феномен выделения корнями целых растений в окружающий раствор стимуляторов дыхания. Физиол. раст., 1995, 42(4): 501-506.
- А н т о н о в А.С. О взаимосвязи геносистематики и геномики. Журн. общ. биол., 2003, 64(2): 181-186.
- О н о С. Генетические механизмы прогрессивной эволюции. М., 1973.
- М а р к а р о в А.М., Г о л о в к о Т.К., Т а б а л е н к о в а Г.Н. Морфофизиология клубнеобразующих растений. СПб, 2001.
- М о к р о н о с о в А.Т. Онтогенетический аспект фотосинтеза. М., 1981.
- Д ь я ч е н к о А.П. Сравнительный анализ структурных и функциональных особенностей фотосинтетического аппарата различных экологических групп высших растений. В сб.: Мезоструктура и функциональная активность фотосинтетического аппарата. Свердловск, 1978: 93-102.
- Г а м а л е й Ю.В. Признаки ксероморфизма. В сб.: Пустынные растения Заалтайской Гоби. Л., 1988: 67-84.
- Ж у р а в л е в а Н.А. Механизм устьичных движений, продукционный процесс и эволюция. Новосибирск, 1992.
Сибирский институт физиологии и биохимии
растений СО РАН,
664033 г. Иркутск, ул. Лермонтова. 132,
e-mail: zhuravleva@sifibr.irk.ru |
Поступила в редакцию
11 мая 2005 года |
SEEDLING-GENETIC MODEL OF HERBACEOUS
FLOWERING PLANT AND THEORY OF SELECTIVE PROCESS
(by the example of potato)
N.A. Zhuravleva
S u m m a r y
The author made an attempt to look at the tuberization theory and the purposeful creation of high productive potato varieties by transgenic methods from more general provisions — as consequence of phylogenetic development of vital form of herbaceous flowering plant, structural unit of which is seedling. Appearance of tuber as the element of seedling is regarding as final stage one of branch of seedling-genetic process in herbaceous dicotyledons, initial special form of which was crown producing seegding and final form — its intensive tuberization form. The model of this process was presented, which permit to explain the origin of main selective potato determinant and give a chance for its control.