СЕЛЬСКОХОЗЯЙСТВЕННАЯ БИОЛОГИЯ, 2008, № 3, с. 48-57
УДК 635.651:577.2:575.17
К ВОПРОСУ О ПРОИСХОЖДЕНИИ ВОЗДЕЛЫВАЕМЫХ БОБОВ И ВНУТРИВИДОВОМ РАЗНООБРАЗИИ Viciafaba L. ПО РЕЗУЛЬТАТАМ МОЛЕКУЛЯРНОГО МАРКИРОВАНИЯ ГЕНОМА
Е.К. ПОТОКИНА, С.В. БУЛЫНЦЕВ, Н. ТОМООКА, Д. ВОГАН
Методом RAPD (random amplified polymorphic DNA ― случайная амплификация полиморфной ДНК) проанализировано генетическое разнообразие образцов местных популяций бобовViciafaba разного географического происхождения, собранных в 1916-1928 годах экспедициями Н.И. Вавилова. Образцы из Северной Африки (Алжир, Египет) продемонстрировали явные генетические отличия от остального внутривидового разнообразия V. faba. Ранее была описана V. pliniana(Trabut) Murat из горных районов Алжира и Западного Марокко, рассматриваемая в качестве возможного предка культурных бобов. Выдвинуто предположение, что генетически обособленные североафриканские популяции V. faba могут представляют собой сравнительно недавно окультуренные формы дикорастущей V. faba, которую ботаники прошлого века описывали как V. pliniana.
Ключевые слова: бобы, внутривидовое разнообразие, RAPD, происхождение культуры.
Key words: faba bean, intraspecific diversity, RAPD, crop origin.
Вопрос о происхождении культурных бобов (Viciafaba L.) — одного из древнейших возделываемых растений Старого Света — остается спорным, несмотря на многочисленные исследования (1-2). Эксперименты с использованием молекулярных маркеров (3-6) не подтвердили близкого родства между V. faba и видами секции Narbonensis, как это прежде предполагалось на основании морфологического сходства. Проведенное нами ранее изучение филогенетических связей 29 видов Vicia с использованием RAPD (random amplified polymorphic DNA) и анализа полиморфизма хлоропластных генов (7) показало, что V. faba при определенной степени родства с ныне существующими видами подрода Vicia не может считаться одомашненной формой одного из них.
Мы попытались определить происхождение культивируемого вида V. faba, проанализировав с помощью молекулярных методов генетическое разнообразие образцов местных популяций бобов разного географического происхождения, собранных экспедициями Н.И. Вавилова, для установления степени генетической близости популяций V.faba из разных участков видового ареала и выявления возможных путей распространения этой культуры.
Методика. Образцы коллекции ВИР (Всероссийский НИИ растениеводства) были собраны в условиях (1916-1928 годы), когда традиционные местные способы ведения сельского хозяйства еще не сменились аграрными технологиями. Систематика и история большинства из них представлены в классической монографии В.С. Муратовой (8). Были проанализированы 46 образцов Viciafaba из коллекции ВИР (собраны и описаны В.С. Муратовой) (8) в сравнении с 3 образцами V. johannisTamam. in Karyag.и 7 образцамиV. narbonensis L. различного географического происхождения. Отметим, что с 1931 года значительную часть образцов из ее коллекции сохранить не удалось, некоторые утратили сходство с оригиналом вследствие переопыления при репродукциях. В результате в опыт были включены только образцы, соответствующие оригиналу. Мы включили в анализ пять образцов, полученных ВИР из Китая, Индии и Пакистана в 1953-1963 годах, чтобы восполнить недостающую информацию.
Растения выращивали на экспериментальном участке NIAR (г. Цукуба, Япония). ДНК выделяли из молодых листьев 8-12 растений каждого образца стандартным методом CTAB (9) со следующими модификациями: 2½CTAB-буфер содержал 1 % 2-меркаптоэтанола, 1 % поливинилпирролидона и 1,4 % метабисульфата натрия. Концентрацию ДНК в пробах доводили до 5 мг/мл с использованием стандартов известной концентрации.
В качестве метода молекулярного маркирования была использована одна из модификаций полимеразной цепной реакции (ПЦР) — RAPD (9). Реакционная смесь для RAPD-ПЦР объемом 10 мкл содержала 10 нг геномной ДНК, 1 мкл 10 PCR-буфера, 0,8 мкл 25 мM MgCl2, 0,2 мкл 10 мM dNTPs, 0,5 мкл 10 мМ праймера, 0,2 мкл Taq-полимеразы в инкубационном буфере (1 ед/мкл). ПЦР проводили в следующем режиме: 1 мин — 94 °С; 30 с — 94 °С, 1 мин — 36 °С и 2 мин — 72 °С (всего 48 циклов); заключительная элонгация — 10 мин 72 °С, после чего пробы охлаждали во льду. Продукты реакции разделяли электрофорезом в 0,8 % агарозном геле в 1½ТВЕ-буфере стандартным способом, окрашивали бромистым этидием и фотодокументировали.
Статистический анализ включал составление суммарной бинарной матрицы для всех праймеров (1 и 0 — соответственно присутствие или отсутствие фрагментов с одинаковой молекулярной массой на электрофореграммах разных образцов). Каждый RAPD-фрагмент рассматривался как отдельный генетический локус. Суммарную матрицу RAPD-спе-ктров для всех видов и образцов подвергали факторному анализу с использованием программного пакета Statistica 5.0. Бинарная матрица, включающая только образцы V. faba, была дополнительно проанализирована методом главных координат (principal coordinates analysis — PCoA) с помощью опций DOUBLE CENTER и EIGENVECTOR программного пакета NTSYS-pc (10).
Результаты. Таксономическая классификация внутривидового разнообразия анализируемых образцов бобов представлена в таблице 1 (по В.С. Муратовой) (8).
Работа выполнена при финансовой поддержке Science and Technology Agency of Japan Fellowships (Япония).
1. Таксономическая характеристика образцов Vicia из коллекции Всероссийского НИИ растениеводства (ВИР), проанализированных методом RAPD-ПЦР |
Образец |
Номер по каталогу ВИР |
Год поступления
в коллекцию |
Происхождение |
Авторство сбора |
Вид, разновидность, форма |
china1 |
51 |
1923, экспедиция |
Китай, Пекин, Калган (Даму) |
В.Е. Писарев |
V.fabavar.equina, f.rosescens |
china2 |
109 |
То же |
Маньчжурия (Дзан-Доу) |
В.Е. Писарев |
V. faba var. major, f. undulosa |
china3 |
1450 |
1953, сельскохозяйственный институт |
Китай (Хан-Чжоу) |
|
V. faba var. major, f.procumbens |
china4 |
1515 |
1953, экспедиция |
Китай, северо-западная провинция, базар |
|
V. faba var. major, f.procumbens |
china5 |
1529 |
1958, сельскохозяйственный институт |
Китай, Ченду, провинция Сычуан, высота 450 м |
|
V.faba |
china6 |
697 |
1927, экспедиция |
Западный Китай, Ханарык |
М.И. Думпис |
V.faba |
spain7 |
802 |
То же |
Испания, Барселона |
Н.И. Вавилов |
V.faba var.equina, f.equina |
spain8 |
788 |
То же |
Испания, Мадрид,
(Робустиано Диез-Обеза) |
Н.И. Вавилов |
V.faba var.equina, f. equina |
spain9 |
924 |
То же |
Испания, Мадрид,
(Робустиано Диез-Обеза) |
Н.И. Вавилов |
V.faba var.equina, f.viridiana |
spain10 |
800 |
То же |
Испания, Мадрид |
Н.И. Вавилов |
V.faba var.minor, f.agrogum |
ethio11 |
741 |
То же |
Эфиопия, Дебарек, 80 км восточнее Гондара, высота 2800-3000 м |
Н.И. Вавилов |
V.faba var.minor, f.abyssinica |
ethio12 |
763 |
То же |
Эфиопия, Дебарек, 80 км восточнее Гондара, высота 2600 м |
Н.И. Вавилов |
V.faba var.minor, f.abyssinica |
Продолжение таблицы 1
|
ethio13 |
773 |
То же |
Эфиопия, провинция Харар, горы Муллат,
высота 2600 м |
Н.И. Вавилов |
V.faba var.minor |
germa14 |
88 |
1922, семенная компания Штрубе |
Германия (Eckendorfer Feldbohnen) |
|
V.faba var.minor, f.agrorum |
franc15 |
266 |
1922, семенная компания Вильморин |
Франция, Париж (Feverole de Picardie) |
|
V.faba var.equina, f.equina |
engla16 |
605 |
1927, семенная компания Суттон |
Англия (Early Mazagan, Broad Bean) |
|
V.faba var.equina, f.equina |
russi17 |
57 |
1923, сельскохозяйственный техникум |
Россия, Костромская обл., Кологривский р-н, местная популяция |
|
V.faba var.minor, f.pumila |
russi18 |
78 |
1922, Пермская селекционная станция |
Россия, Пермская обл. |
|
V.faba var.equina, f.opaca |
russi19 |
108 |
1922, Уральская селекционная станция |
Россия, Урал, местная популяция |
|
V.faba var.minor, f.borealis |
russi20 |
119 |
1922, сельскохозяйственный техникум |
Россия, Курская обл., местная популяция |
|
V. faba var. minor, f.agrorum |
russi21 |
185 |
1922, сельскохозяйственный техникум |
Россия, Тверская обл., местная популяция |
|
V. faba var. major, f.ianthina |
russi22 |
547 |
1927, Волынская селекционная станция |
Украина, Винницкая обл. |
|
V. faba var. minor, f.violacea |
syria23 |
478 |
1926, экспедиция |
Сирия, Дамаск |
Н.И. Вавилов |
V. faba var. equina, f.mediterranea |
italy24 |
630 |
1927, экспедиция |
Италия, Рим (Favette) |
Н.И. Вавилов |
V.faba var.equina, f.leucosperma |
italy25 |
634 |
То же |
Италия, Рим |
Н.И. Вавилов |
V.faba var.equina, f.equina |
italy26 |
942 |
То же |
Италия, Милан (Fave) |
Н.И. Вавилов |
V.faba var.equina, f.leucosperma |
sardi27 |
851 |
То же |
Сардиния, Сассари |
Н.И. Вавилов |
V.faba var.equina, f.divulgata |
india28 |
1099 |
1928, экспедиция |
Индия, Кашмир (бокля) |
В.В. Маркович |
V.faba var.minor |
india29 |
1102 |
То же |
Индия, Кашмир (бокля) |
В.В. Маркович |
V.faba var.minor |
india30 |
1559 |
1959, посольство |
Индия, Уттар-Прадеш |
|
V. faba ssp.paucijugaf.bacla |
pakis31 |
2182 |
1963, экспедиция |
Пакистан |
|
V. faba ssp.paucijugaf.bacla |
afgan32 |
161 |
1917, экспедиция |
Афганистан, Памир,
Ванч |
Н.И. Вавилов |
V.faba var.minor f.asiatica |
afgan33 |
209 |
1924, экспедиция |
Афганистан, Герат,
базар |
Н.И. Вавилов |
V. faba var. minor f.farachica |
afgan34 |
215 |
То же |
Афганистан, Кабул, Сари-Чешме |
Н.И. Вавилов |
V.faba var.minor f.nigra |
afgan35 |
225 |
То же |
Афганистан, Бамиан |
Н.И. Вавилов |
V.faba var.minor f.laterita |
afgan36 |
238 |
То же |
Афганистан, Кабул |
Н.И. Вавилов |
V.faba var.minor f.laterita |
alger37 |
490 |
1926, экспедиция |
Алжир, Тиарет |
Н.И. Вавилов |
V. faba var. equina, f.mediterranea |
egypt38 |
872 |
1927, экспедиция |
Египет, Медио Эгитто, Каир |
Н.И. Вавилов |
V.faba var.minor f.aegyptica |
egypt39 |
883 |
То же |
Египет, Альто Эгитто |
Н.И. Вавилов |
V.faba var.minor f.aegyptica |
turke40 |
149 |
1918, экспедиция |
Туркестан (Туркестанские бобы) |
Н.И. Вавилов |
V.faba var.minor f.asiatica |
daghe41 |
144 |
1917, экспедиция |
Дагестан, Гуниб |
Н.И. Вавилов |
V. faba var. minor f.daghestanica |
daghe42 |
283 |
1925, экспедиция |
Дагестан, Андинский округ |
|
V. faba var. minor f.daghestanica |
armen43 |
186 |
1917, экспедиция |
Армения, Дегал-оглы, высота 4600 м |
|
V. aba var. major
f.orbicularis,
f.megalosperma |
armen44 |
281 |
1925, экспедиция |
Армения, Гори |
Е.И. Барулина |
V.faba var.major
f.megalosperma |
georg45 |
987 |
1927, экспедиция |
Грузия, Верхняя Сванетия |
|
V. faba var. minor f.svanetica |
georg46 |
1407 |
1948, экспедиция |
Грузия, Верхняя Сванетия |
В.С. Муратова |
V. fabavar.minor |
johan47 |
36718 |
ВИР |
Азербайджан |
|
V. johannis |
johan48 |
35387 |
То же |
Туркмения |
|
V. johannis |
johan49 |
33191 |
То же |
Туркмения |
|
V. johannis |
narbo50 |
35225 |
То же |
Турция |
|
V.narbonensis |
narbo51 |
35224 |
То же |
Грузия |
|
V.narbonensis |
narbo52 |
35392 |
То же |
Грузия |
|
V.narbonensis |
Продолжение таблицы 1 |
narbo53 |
34415 |
То же |
Португалия |
|
V.narbonensis |
narbo54 |
34827 |
То же |
Казахстан |
|
V.narbonensis |
narbo55 |
35162 |
То же |
Турция |
|
V.narbonensis |
narbo56 |
35089 |
То же |
Сирия |
|
V.narbonensis |
П р и м е ч а н и е. Местные названия бобов указаны в скобках. Пропуски означают отсутствие информации об авторстве сборов. |
Декамерные олигонуклеотидные праймеры (табл. 2), использованные для анализа 46 образцов V. faba, 3 образцов V. johannis и 7 образцов V. narbonensis, амплифицировали 246 хорошо различимых RAPD-фрагментов (каждый праймер — от 6 до 25 фрагментов на один образец).
2. Нуклеотидные последовательности декамерных олигонуклеотидных праймеров, использованных для RAPD-ПЦР-анализа |
Праймер |
Сиквенс праймера |
P7 |
3´-AGCACTTCGG-5´ |
P9 |
3´-ACTCCGCAGT-5´ |
P19 |
3´-TAGACAGTCG-5´ |
P22 |
3´-ATGAGTCCAC-5´ |
P54 |
3´-CGTAGCGCGA-5´ |
P68 |
3´-CATAGGCCCT-5´ |
P105 |
3´-TGGTCGCTGA-5´ |
P120 |
3´-TGGTCACAGA-5´ |
P152 |
3´-GTTTCGCTCC-5´ |
P155 |
3´-GGACTGGAGT-5´ |
P158 |
3´-GGTGACGCAG-5´ |
P163 |
3´-CCTTGACGCA-5´ |
P195 |
3´-GTAGGCGTCG-5´ |
P205 |
3´-GCCGTGAAGT-5´ |
P206 |
3´-CTCGACTAGG-5´ |
P226 |
3´-CCAGACAAGC-5´ |
P284 |
3´-TCCATGCCGT-5´ |
P300 |
3´-CAGGCGGCGT-5´ |
P301 |
3´-GCTGGACATC-5´ |
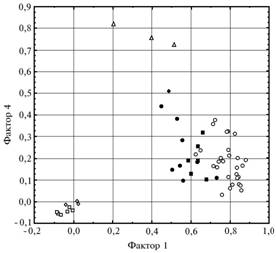
С помощью факторного анализа, выявляющего структуру бинарной матрицы RAPD-профи-лей и коррелирующие переменные (которые, следовательно, детерминируются общим фактором), было обнаружено шесть главных факторов в структуре корреляций у полученных RAPD-спектров (табл. 3). На основании максимальной корреляции RAPD-профиля и соответствующего фактора (фактор-ной нагрузки) изученные образцы разделялись на 6 групп. Так, три первых и наиболее существенных фактора определялись меж-видовой изменчивостью: первый обособляет образцы V. faba, второй — V. narbonensis и третий — V. johannis.Остальные три фактора характеризовали структуру внутривидовой изменчивости V. faba. Среди них наиболее весомый компонент изменчивости — четвертый фактор был связан с образцами североафриканского происхождения (см. табл. 3, группа 4), пятый — обособлял образцы из Афганистана, Пакистана и Индии, шестой — объединял группу так называемых черных русских бобов. Структура генетического разнообразия V. faba и близкородственных видов по результатам RAPD-анализа в двухмерном пространстве, заданном первым и четвертым факторами, приведена на рисунке 1.
Дополнительную информацию о структуре внутривидового генетического разнообразия староместных популяций бобов получили при анализе суммарной матрицы попарного сходства RAPD-спектров образцов V. faba (рис. 2) методом главных координат (PCoA), известным также как метрическое шкалирование, позволяющее отобразить корреляцию между RAPD-спектрами анализируемых образцов в системе двух координат, с которыми связан наибольший процент наблюдаемой изменчивости. Результаты метрического шкалирования подтвердили, что изучаемые популяции бобов (за исключением образцов из Алжира и Египта) генетически весьма близки, что соответствует результатам более ранних исследований (1, 11, 12). С помощью RAPD-анализа также не удалось выявить различия между ботаническими разновидностями (varieties, далее в тексте обозначаемые какvar.)и формами (formae, далееf.). Большинство образцов объединялось в центральную группу средиземноморского и европейского происхождения (см. рис. 2, I). Близкими к центральной группе были популяции Кавказа, южной России и Ближнего Востока (см. рис. 2, I, а; I, б). Дивергентную группу образовали популяции из Индии и Афганистана (см. рис. 2, II, III), которые демонстрировали также явное сходство с образцами черных русских бобов (см. рис. 2, IV). Наиболее генетически обособленными оказались североафриканские популяции из Алжира и Египта (см. рис. 2, V).
Полученные результаты послужили основанием для обсуждения гипотез о происхождении и географическом распространении культуры бобов.
Индия. По мнению В.С. Муратовой (8), культура бобов в Индии не является древней. Тем не менее, автор указывает на некоторые эндемичные формы из районов Lyullpur, Indore и Pusa и описывает их в ранге подвида V. faba ssp. рaucijuga Murat. В нашем исследовании указанный таксон был представлен двумя образцами (india30 и pakis31, см. рис. 2), которые вместе с тремя другими из Афганистана обособляются от остальных популяций V. fabaна графике главных координат (см. рис. 2, II). Образцы из Кашмира, собранные В.В. Марковичем в 1928 году (india28 и india29) и классифицированные В.С. Муратовой как аналогичные образцам из Эфиопии (var. minor и f. abyssinica) (см. рис. 2, I), тяготели к центральной группе, что указывает на их высокое генетическое сходство со средиземноморскими и европейскими популяциями.
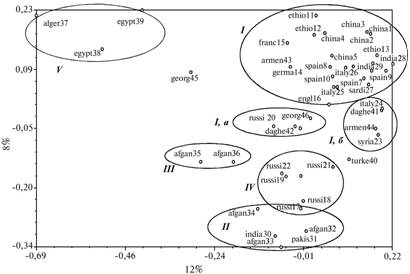 |
Рис. 2. Структура внутривидовой изменчивости RAPD-спектров у образцов V. faba, выявленная методом главных координат. I-V — группы происхождения, отражающие корреляцию RAPD-спектров. |
Афганистан. Особенность афганских образцов бобов, собранных Н.И. Вавиловым в 1924 году, — эндемизм и разнообразие форм (8). Для них характерно варьирование размера семян (от мелких до средних) и окраски семенной кожуры (розовая, зеленая, черная). Четыре ботанические формы, выделенные В.С. Муратовой из этих популяций, мы проанализировали RAPD-методом. Из них три — f. asiatica, f. farachica, f. nigra(afgan32, afgan33, afgan34, см. рис. 2, II) показали генетическое сходство с образцами V. faba subsp. paucijuga из Индии и Пакистана. Генетически удаленными от ssp. paucijuga оказались афганские образцы, описанные как f. laterita (afgan35, afgan36, см. рис. 2, III) из ущелий Бамиана и Кабула, где в начале прошлого века бобы были второй по значимости возделываемой культурой после пшеницы (8). Наши результаты показывают, что популяции бобов из Индии и Пакистана и частично Афганистана генетически близки друг к другу и обособляются от образцов из Европы, Западного Средиземноморья и Ближнего Востока. По результатам RAPD-анализа, описанные ботанические разновидности и формы из этих районов неотличимы на уровне ДНК.
3. Факторные нагрузки главных компонент изменчивости у изученных образцов Vicia по результатам RAPD-ПЦР-анализа |
Образец |
Группа |
Фактор |
1 |
2 |
3 |
4 |
5 |
6 |
china1 |
1 |
0,881 |
0,012 |
0,016 |
0,193 |
0,105 |
0,033 |
china2 |
1 |
0,790 |
0,037 |
-0,007 |
0,325 |
0,164 |
-0,041 |
china3 |
1 |
0,835 |
0,047 |
0,039 |
0,158 |
0,059 |
0,049 |
china4 |
1 |
0,797 |
0,029 |
0,017 |
0,213 |
0,212 |
0,015 |
china5 |
1 |
0,793 |
0,077 |
-0,032 |
0,239 |
0,061 |
0,208 |
china6 |
1 |
0,866 |
-0,007 |
0,013 |
0,165 |
0,172 |
0,119 |
spain7 |
1 |
0,793 |
0,038 |
0,025 |
0,115 |
0,141 |
0,120 |
spain8 |
1 |
0,743 |
0,073 |
-0,028 |
0,186 |
0,058 |
0,188 |
spain9 |
1 |
0,843 |
0,039 |
0,006 |
0,114 |
0,110 |
0,059 |
spain10 |
1 |
0,735 |
0,095 |
-0,037 |
0,157 |
0,125 |
0,167 |
ethio11 |
1 |
0,838 |
0.037 |
0,002 |
0,109 |
0,079 |
0,155 |
ethio12 |
1 |
0,782 |
0,008 |
0,019 |
0,323 |
0,001 |
0,160 |
ethio13 |
1 |
0,827 |
0,012 |
0,009 |
0,315 |
0,102 |
0,102 |
germa14 |
1 |
0,753 |
0,001 |
0,014 |
0,202 |
0,146 |
0,152 |
franc15 |
1 |
0,726 |
-0,008 |
0,047 |
0,376 |
0,140 |
0,102 |
engla16 |
1 |
0,717 |
0,111 |
-0,074 |
0,163 |
0,163 |
0,181 |
russi17 |
6 |
0,601 |
0,093 |
-0,106 |
0,129 |
0,262 |
0,543 |
russi18 |
6 |
0,636 |
0,029 |
-0,034 |
0,187 |
0,299 |
0,380 |
russi19 |
6 |
0,589 |
0,061 |
-0,090 |
0,191 |
0,195 |
0,552 |
russi20 |
6 |
0,663 |
0,075 |
-0,025 |
0,318 |
0,150 |
0,496 |
russi21 |
6 |
0,681 |
0,046 |
-0,081 |
0,103 |
0,207 |
0,484 |
russi22 |
6 |
0,635 |
0,093 |
-0,008 |
0,256 |
0,318 |
0,422 |
syria23 |
1 |
0,801 |
0,005 |
0,009 |
0,062 |
0,361 |
0,054 |
italy24 |
1 |
0,846 |
0,042 |
-0,023 |
0,080 |
0,236 |
0,150 |
italy25 |
1 |
0,764 |
0,065 |
-0,032 |
0,187 |
0,144 |
0,216 |
italy26 |
1 |
0,842 |
0,014 |
0,024 |
0,202 |
0,113 |
0,219 |
sardi27 |
1 |
0,835 |
0,048 |
-0,004 |
0,129 |
0,206 |
0,075 |
india28 |
1 |
0,860 |
0,036 |
0,008 |
0,054 |
0,093 |
0,109 |
india29 |
1 |
0,854 |
0,054 |
0,014 |
0,077 |
0,125 |
0,109 |
india30 |
5 |
0,506 |
0.119 |
-0,002 |
0,148 |
0,489 |
0,329 |
pakis31 |
5 |
0,561 |
0,038 |
0,030 |
0,098 |
0,518 |
0,276 |
afgan32 |
5 |
0,635 |
0,103 |
-0,024 |
0,185 |
0,538 |
0,154 |
afgan33 |
5 |
0,544 |
0,104 |
-0,070 |
0.167 |
0,655 |
0,093 |
afgan34 |
5 |
0,557 |
0,070 |
-0,007 |
0,283 |
0,560 |
0,163 |
afgan35 |
5 |
0,448 |
0,064 |
-0,071 |
0,440 |
0,403 |
0,209 |
afgan36 |
5 |
0,529 |
0,057 |
-0,102 |
0,383 |
0,452 |
0,113 |
algyr_37 |
4 |
0,205 |
0,066 |
-0,001 |
0,818 |
0,087 |
0,011 |
egypt_38 |
4 |
0,400 |
0,123 |
-0,035 |
0.755 |
0,154 |
0,157 |
egypt_39 |
4 |
0,514 |
0,093 |
0,020 |
0,723 |
0,079 |
0,107 |
turke40 |
5 |
0,730 |
0,010 |
-0,079 |
0,112 |
0,456 |
-0,001 |
daghe41 |
1 |
0,813 |
-0,012 |
-0,056 |
0,079 |
0,360 |
-0,121 |
daghe42 |
1 |
0,624 |
0,028 |
-0,075 |
0,218 |
0,335 |
0,022 |
armen43 |
1 |
0,713 |
0,023 |
-0,009 |
0,357 |
0,239 |
0,016 |
armen44 |
1 |
0,757 |
0,013 |
-0,044 |
0,033 |
0,319 |
0,005 |
georg45 |
7 |
0,488 |
0,089 |
0,019 |
0,511 |
0,187 |
0,116 |
georg46 |
1 |
0,649 |
0,026 |
0,012 |
0,236 |
0,211 |
0,314 |
Продолжение таблицы 3
|
johan47 |
3 |
0,024 |
-0,273 |
0,923 |
-0,008 |
-0,039 |
-0,039 |
italy48 |
3 |
-0,042 |
-0,227 |
0,941 |
-0,016 |
-0,006 |
-0,004 |
italy49 |
3 |
0,016 |
-0,235 |
0,930 |
0,001 |
-0,042 |
-0,044 |
narbo50 |
2 |
-0,032 |
-0,962 |
0,132 |
-0,047 |
-0,042 |
-0,015 |
narbo51 |
2 |
-0,004 |
-0,959 |
0,143 |
-0,042 |
-0,063 |
-0,046 |
narbo52 |
2 |
-0,065 |
-0,964 |
0,101 |
-0,063 |
-0,023 |
-0,020 |
narbo53 |
2 |
-0,082 |
-0,937 |
0,086 |
-0,056 |
0,009 |
0,013 |
narbo54 |
2 |
-0,086 |
-0,947 |
0,075 |
-0,048 |
-0,002 |
-0,012 |
narbo55 |
2 |
-0,004 |
-0,963 |
0,111 |
-0,041 |
-0,064 |
-0,047 |
narbo56 |
2 |
-0,023 |
-0,937 |
0,119 |
-0,026 |
-0,045 |
-0,040 |
Общая дисперсия |
23,87 |
6,77 |
2,78 |
4,03 |
3,01 |
2,76 |
Доля общей дисперсии |
0,42 |
0,12 |
0,05 |
0,07 |
0,05 |
0,05 |
П р и м е ч а н и е. Подчеркнуты максимальные факторные нагрузки (показатели корреляции RAPD-профиля). |
Кавказ. При анализе шести образцов из Армении, Дагестана и Грузии два образца из Армении, определенные В.С. Муратовой как var. major (f. orbicularis и f. megalosperma) группировались вместе со средиземноморскими популяциями, где указанная разновидность встречается повсеместно. Один из двух дагестанских образцов, отнесенных к разновидности var. minor f. daghestanica(daghe41), находился в кластере образцов из Италии и Сирии (см. рис. 2, I, б). Это подтверждает гипотезу В.С. Муратовой о близости кавказских и средиземноморских бобов. Второй из дагестанских образцов (daghe42) показал генетическое сходство с образцом из южных районов России, определенным как var. minor f. agrorum. Эта разновидность широко распространена среди так называемых германских кормовых бобов и, по В.С. Муратовой, была интродуцирована из Европы в западные районы России несколько столетий назад.
В.С. Муратовой также были описаны несколько эндемичных кавказских форм V. faba(8). Одна из них (var. minor f. svanetica) из верхней Сванетии выделена автором на основании внешнего сходства с германскими кормовыми бобами (f. agrorum)по специфическому габитусу пригораздо более раннем созревании. В наших исследованиях один образец из верхней Сванетии (georg46), идентифицированный В.С. Муратовой как f. svanetica, действительно оказался очень сходным по RAPD-профилю с образцом f. agrorum из России (см. рис. 2, I, a), другой (georg45, также f. svanetica) занимал особое место на графике главных координат, будучи значительно обособленным от всех остальных кавказских образцов. Полученные результаты RAPD-анализа свидетельствуют в пользу предположения В.С. Муратовой о том, что в Закавказье представлены формы бобов, интродуцированные в разное время как с востока (южные районы России), так и с запада (европейские страны), а также что некоторые высокогорные районы характеризуются эндемичными формами (8).
Европейская часть России и Сибирь. При изучении шести образцов с территории России образец russi20 (см. рис. 2) из Курской области был отнесен к f. agrorum, остальные пять — к группе черных русских бобов, названных так из-за преобладающей темно-фиолетовой окраски семенной кожуры. Черные русские бобы возделываются в северных и центральных районах России, а также на южном Алтае, в Иркутской области и Забайкалье и объединяют несколько ботанических разновидностей и форм: var. minor (f. pumilaи f. borealis), var. equina f. opaca, var. major f. ianthina. В.С. Муратова отмечает, что у северных границ видового ареала преимущественно распространены формы с темной семенной кожурой, в южных и центральных областях России — со светлой. По мнению автора, светлосемянные формы f. orbicularis и f. megalosperma, а также мелкосемянные f. agrorum были интродуцированы в западные, центральные и южные районы России из Европы. Единственный светлосемянный образец f. agrorum из Курска в нашем исследовании (russi20) действительно группируется вместе с евро-средиземноморскими популяциями (см. рис. 2). Черные русские бобы (russi17, 18, 19, 21, 22) проявляли явное генетическое сходство с образцами из Индии и Афганистана, то есть по результатам RAPD-анализа они могут иметь азиатское происхождение. Это противоречит гипотезе В.С. Муратовой об интродукции черных русских бобов болгарскими огородниками, которые в XI-XII веке начали заселять южные районы России.
Китай. Пять образцов из Китая (china1, china2, china3, china4, china5) (см. рис. 2) группировались вместе с образцами западного Средиземноморья, Эфиопии и Центральной Европы. Этот факт подтверждает гипотезу о возможности распространения культуры бобов из средиземноморских стран в Китай по Великому шелковому пути (13).
Эфиопия.У образцов V. faba, собранных экспедицией Н.И. Вавилова в 1927 году в Абиссинии на высоте 1800-2800 м, обнаружили тесную связь с центральной группой средиземноморского происхождения. Таким образом, RAPD-анализ не выявил существенной дивергенции эфиопских популяций, как это прежде предполагалось на основе морфологических (8) и биохимических (12) данных.
Побережье Северной Африки. Образец из Алжира (окрестности г. Ти-арет) морфологически был идентифицирован как f. mediterranea, два других образца из Египта — как var. minor f. aegyptiaca. Последняя ботаническая форма, по В.С. Муратовой, имеет очень ограниченное распространение и характеризуется мелкими сплюснутыми семенами, выделяющими египетские образцы из всего разнообразия форм, представленных на североафриканском побережье. В наших опытах все три образца (независимо от принадлежности к ботанической разновидности и форме) на уровне ДНК демонстрировали наиболее явную генетическую дивергенцию от остального внутривидового разнообразия возделываемой культуры V. faba.
Происхождение и распространение культуры бобов. Обнаруженная нами генетическая обособленность североафриканских популяций культурных бобов, по своей амплитуде сопоставимая с межвидовой изменчивостью, позволяет переосмыслить существующие гипотезы о предковой форме культурного вида V. faba. Северная Африка имеет особое отношение к обсуждаемому вопросу, так как в 1893 году в горах Алжира (at Sersou de Boubaraki a Vialar) на высоте 900 метров французским ботаником L. Trabut в дикорастущем состоянии была описана популяция бобов, существенно отличающихся от типичной V. faba. По свидетельству автора, африканские женщины собирали зеленые плоды дикорастущих бобов для приготовления пищи (14, цит. по 8). L. Trabut описал найденные растения в подвидовом ранге как FabavulgarisPliniana Trabut, полагая, что морфологические особенности этих растений недостаточно выражены для выделения нового вида. В.С. Муратова (8), изучив морфологические признаки типового экземпляра и анатомическую структуру семенной кожуры, а также учитывая дикорастущее состояние формы, выделила ее как самостоятельный вид V. pliniana(Trabut) Murat, впервые предложив рассматривать его в качестве возможного предка культурных бобов.
Известны и другие сообщения об обнаружении V. pliniana в Северной Африке в начале прошлого века. Так, R. Maire (15) описывает растение, отличающееся от V. faba более короткими (до 45 мм) трехсемянными бобами. Оно было найдено в западной части Марокко (Capar-Boulhaut) в дикорастущем состоянии на кварцитовых отложениях, пастбищах и около кустарников. Среди образцов V. pliniana из гербария Монпелье (Herbier Battandier, MPU, Montpellir) есть экземпляры, собранные в Алжире в местечке at Sersou de Tiaret. Именно из этого географического пункта происходит образец, отобранный Н.И. Вавиловым в 1927 году и включенный в наше исследование (alger37, см. рис. 2, V). Приняв во внимание гипотезу J.I. Cubero (16), согласно которой не удается выявить дикорастущие формы бобов, поскольку в районах произрастания они непосредственно переводятся человеком в культивируемое состояние, предполагаемой причиной явной генетической обособленности североафриканских популяций V. faba можно считать то, что они представляют собой сравнительно недавно окультуренные формы дикорастущей V. faba, которую ботаники прошлого века описывали как V. pliniana.
Таким образом, результаты нашего исследования показывают, что дикорастущий предок культурных бобов, очень незначительно отличающийся морфологически от современных форм V. faba, в историческое время мог быть широко распространен в Средиземноморье и на Ближнем Востоке, как практически все другие виды подрода Vicia. По данным N. Maxted (17), наивысшая концентрация видового разнообразия и возможный центр происхождения подрода Vicia приходится на юго-восточные регионы Европы и Юго-Западную Азию. Оттуда виды распространялись на север и запад, в меньшей степени — на восток и юг. То есть представляется вполне возможным, что дикорастущая форма V. fabaдействительно была найдена в начале прошлого столетия в горах североафриканского побережья.
Наши результаты хорошо согласуются с гипотезой D. Zohary и M. Hopf (18) о том, что возделывание и использование бобов началось на Ближнем Востоке, и затем эта культура широко распространилась во времена неолита. Это не означает, однако, что бобы не были известны в других районах Средиземноморья (16): их название встречается в четырех древних языках — греческом, арабском, древнееврейском и языке берберов (8, 13). Возможно, дикорастущий предок бобов в то время был распространен в пределах своего естественного ареала, откуда происходят и другие возделываемые виды Vicia.
Пути распространения культуры бобов. J.I. Cubero (16) высказал предположение о четырех основных направлениях распространения бобов из стран Ближнего Востока, с чем полностью согласуются результаты нашего исследования. Районы Средиземноморья и Ближнего Востока, по-видимому, представляют собой первичный центр происхождения бобов. Подобно другим видам Vicia, бобы далее распространились в Среднюю Азию, Афганистан и Индию, где сформировался вторичный центр формового разнообразия. Из Средиземноморья бобы распространились также в европейские страны, образовав третичный центр разнообразия (8).
Итак, проведенный RAPD-анализ не выявил явной генетической дифференциации эфиопских образцов от популяций средиземноморского ядра. Образцы из Китая на уровне ДНК, как оказалось, также практически неотличимы от средиземноморских и европейских форм. То же можно сказать о популяциях с Кавказа, куда бобы могли быть интродуцированы русскими поселенцами. В Россию бобы проникли, очевидно, из Европы (за исключением черных русских бобов, которые, по результатам нашего анализа, имеют азиатское происхождение). Видовое и формовое разнообразие Vicia в горах Алжира и Западного Марокко заслуживает специального исследования для получения более полной информации о таксоне V. pliniana, который может иметь непосредственное отношение к дикорастущим предковым формам культивируемых бобов.
Авторы выражают благодарность Л.Л. Малышеву (ВИР) за помощь в статистической обработке результатов.
Л И Т Е Р А Т У Р А
- L a d i z i n s k y G. Seed protein electrophoresis of the wild and cultivated species of section Faba of Vicia. Euphytica, 1975, 24: 785-788.
- M a x t e d N., K h a t t a b A.M.A., B i s b y F.A. The newly discovered relatives of Vicia faba L. do little to resolve the enigma of its origin. Bot. Chron., 1991, 10: 435-465.
- V a n de V e n W.T.G., D u n c a n N., R a m s e y G. e.a. Genetic variation and systematic relationships in Vicia. In: Advances in Legume systematic. 8. Legumes of economic importance /Eds. B. Pickersgill, J.M. Lock. Kew, 1996: 31-40.
- P r z y b y l s k a J., Z i m n i a k - P r z y b y l s k a Z. Electrophoretic seed albumin patterns and species relationship in Vicia sect. Faba (Fabaceae). Pl. Syst. Evol., 1995, 198: 179-194.
- R a i n a S.N., O g i h a r a Y. Chloroplast DNA diversity in Vicia faba and its close relatives: implications and reassessment. Theor. Appl. Genet., 1994, 88: 261-266.
- J a a s k a V. Isoenzyme diversity and phylogenetic affinities in Vicia subgenus Vicia (Faba-ceae). Genet. Res. Crop Evol., 1997, 44: 557-574.
- P o t o k i n a E., T o m o o k a N., V a u g h a n D.A. e.a. Phylogeny of Vicia subgenus Vicia (Fabaceae) based on analysis of RAPDs and RFLP of PCR-amplified chloroplast genes. Genet. Res. Crop Evol., 1999, 46(2): 149-161.
- М у р а т о в а В.С. Бобы (Vicia faba L.). Тр. по прикл. бот., ген. и сел., 1931, 50: 1-285.
- W i l l i a m s J.G., H a n a f e y M.K., R a f a l s k i J.A. e.a. Genetic analysis using randomly amplified polymorphic DNA markers. Methods Enzymol., 1993, 218: 704-740.
- R h o l f F.J. NTSYS-pc: numerical taxonomy and multivariate analysis system. N.-Y., 1992.
- P o l i g n a n o G.B., S o l e n d i d o R., U g g e n t i R. Protein polymorphism among genotypes of faba bean from Afganistan and Ethiopia. Fabis Newslet., 1991, 28-29: 8-11.
- S e r a d i l l a J.M., M o r a T.D., M o r e n o M.T. Geographic dispersion and varietal diversity in Vicia faba L. Genet. Res. Crop Evol., 1993, 40: 143-151.
- H a n e l t P. Zur Geschichte des Anbaues von Vicia faba L. und ihrer verschidenen Formen. Die Kulturpflanze, 1972, 20: 75-128.
- T r a b u t L. L’indigenat de la Flore en Algerie. Bul. De la Soc. Nat. De l’Afrique, 1911, 7-1 Juillet: 1-7.
- M a i r e R. Contribution a l’etude de la flore l’Afrique du Nord. Bul. De le Soc. D’Hist. Nat. de l’Afrique. 1929, 20.
- C u b e r o J.I. On the evolution of Vicia faba L. Theor. Appl. Genet., 1974, 45: 47-51.
- M a x t e d N. An ecogeographical study of Vicia subgenus Vicia. Systematic and Ecogeo-graphical studies on crop genepools. V. 8. Rome, 1995.
- Z o h a r y D., H o p f M. Domestication of pulses in the Old World. Science, 1973, 182: 887-894.
ГНУ ГНЦ РФ Всероссийский НИИ
растениеводства им. Н.И. Вавилова,
190000 г. С.-Петербург, ул. Большая Морская, 44,
e-mail: e.potokina@vir.nw.ru, e.potokina@yahoo.com;
Генбанк, Национальныйинститутагробиологическихнаук (National Institute of Agrobiological Resources ―NIAR),
Kannondai 2-1-2, Tsukuba, Ibaraki, 305, Japan (г. Цукуба, Япония),
e-mail: duncan@abr.affrc.go.jp |
Поступила в редакцию
29 декабря 2007 года |
TO THE QUESTION ABOUT ORIGIN OF CULTIVATED BEAN
AND INTRASPECIFIC VARIABILITY IN Viciafaba L. ACCORDING
TO THE RESULTS OF MOLECULAR MARKING OF GENOME
E.K. Potokina, S.V. Bulyntsev, N. Tomooka, D. Vogan
S u m m a r y
By the method of RAPD the authors analyzed the genetic variability of local populations of Vicia faba of different geographic origin collected in 1916-1928 years during N.I. Vavilov’s expeditions. The specimens from North Africa (Algeria, Egypt) have sharp genetic distinctions from remaining V. faba intraspecific variability. V. pliniana(Trabut) Murat, described before, from mountain areas of Algeria and West Marocco is considered as possible ancestor of cultural beans. It was supposed that genetic isolated North African populations of V. faba are relatively recently tame forms of wild V. faba, which was described by botanists of last century as V.pliniana.