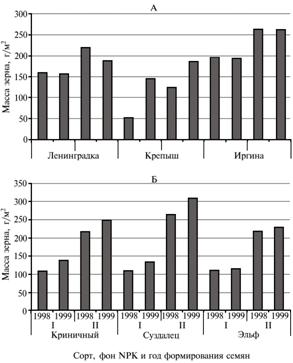
Рис. 1. Урожайность зерна у сортов пшеницы (А) и ячменя (Б) в зависимости от года формирования семян и фона NPK. Варианты I и II — соответственно без внесения и с внесением удобрений.
Проблемы адаптивности зерновых культур
УДК 633.111.1+633.162]:631.524.85:581.5
Адаптивность злаков (Poaceae) в связи с условиями
превегетации и вегетации
Н.А. ЛЫКОВА
На трех сортах яровой пшеницы и трех сортах ярового ячменя в полевых опытах на фоне действия разных почвенно-климатических и эколого-географических факторов изучали влияние условий формирования семян на их продуктивные и адаптивные свойства в период вегетации потомственных растений. Выявлены различия по фенотипическим эффектам между видами TriticumaestivumL. и HordeumvulgareL., а также между сортами внутри одного вида. Показано, что онтогенетическую адаптивность пшеницы изменяли условия превегетации во время формирования двух последних метамеров (7-го и 8-го снизу). Контроль за гидротермическими факторами при выращивании растений материнского поколения снижал риск искажения результатов эколого-генетического испытания однолетних злаков.
Ключевые слова: факторы среды, последействия, Poaceae, морфогенез, адаптивность.
Key words: environmental factors, aftereffects, Poaceae, morphgenesis, adaptability.
Адаптивность как способность сохранять жизнеспособность и формировать семена в условиях, отличных от оптимальных (вплоть до экстремальных), — важное свойство растений. Продуктивность растений во многом зависит от внешних факторов, в частности минерального и органического питания, гидротермических условий вегетации. Изучение прохождения фаз морфогенеза растений и формирования жизнеспособных семян в широком диапазоне условий произрастания позволяет выявить фенотипическую реакцию сорта на условия среды и является предметом эколого-генетического испытания новых сортов и гибридов. Изменение этой реакции в онтогенезе при рассмотрении последовательно морфогенетических признаков, формирующихся в разные фазы развития, показывает онтогенетическую адаптивность растений.
Экологические (экофизиологические) последействия условий среды, в частности питания растений (1, 2), температуры (3) и сроков посева (4), установлены для фенотипа потомственных растений, особенно при воздействии лимитирующих факторов. Экологические последействия обычно изучают у проростков и ювенильных растений (2, 5, 6). Исследования влияния материнских фенотипических эффектов на потомственные растения последующих репродукций немногочисленны (7). Определена видо- и сортоспецифичность материнского фенотипического эффекта (8, 9).
Вегетацией обычно называют жизнедеятельность растений потомственного поколения. Для определения жизненного цикла семенных растений нами предложен термин «превегетация». Отсутствием единой точки зрения на степень и механизмы влияния условий превегетации на адаптивность вегетирующих растений обусловлена актуальность подобных исследований.
Целью представленной работы было изучение закономерностей проявления материнского фенотипического эффекта в модельном эксперименте с однолетними злаками, в частности определение границ условий превегетации, при которых могут быть сформированы однотипные семена, и особенностей формирования адаптивности в онтогенезе растений автогамных видов злаков.
Методика. Объектом исследования служили семена первой репродукции сортов яровой пшеницы Ленинградка, Крепыш, Иргина (TriticumaestivumL.) и ярового ячменя Криничный, Суздалец, Эльф(Hordeum vulgare L.). Сортовая чистота семян пшеницы подтверждена гель-элек-трофорезом глютенинов после вегетации 2000 года.
Опыты проводили в течение трех лет: 1998 или 1999 год — превегетация растений, 2000 — вегетация (собственно эксперимент). Семена, полученные в 1998 и в 1999 годах, высевали в 2000 году по схеме: 1998®2000 и 1999®2000.
В 1998-1999 годы растения выращивали в Гатчинском районе Ленинградской области на Меньковской опытной станции Агрофизического НИИ (59°34'N, 30°13'E). Почва дерново-слабоподзолистая глееватая. В 1998 и 1999 году содержание в пахотном слое подвижных форм фосфора — соответственно 8,2 и 4,0 мг/100 г почвы, калия — 4,4 и 1,0 мг/100 г почвы; внесение минерального азота 3-кратное — соответственно 12,2; 27,2; 48,8 и 8,0; 14,0; 20,0 мг/100 г почвы. Посев проводили сеялкой СМ-16 на делянках площадью 5 м2 из расчета 500 всхожих семян на 1 м2. Запасы доступной для растений влаги в метровом слое почвы в течение вегетационного периода характеризовались тремя уровнями каждый год: в 1998 году — 311, 457, 546 мм, в 1999 — 210, 249, 378 мм.
Гидротермический коэффициент (ГТК) рассчитывали по формуле: ГТК = 10r/t, где r — сумма осадков (мм), t — сумма среднесуточных температур выше 10 °C (показатели за вегетацию). Он соответствовал содержанию запасов доступной влаги в метровом слое за вегетационный период и был равен в 1998 и 1999 году соответственно 2,0; 2,9; 3,4 и 1,4; 1,7; 2,6. Полученные семена хранили в жестяных банках при температуре 20 °С и влажности воздуха 70 %.
В 2000 году эксперимент проводили в Курской области (51°48'N, 36°06'E). Почва — типичный чернозем тяжелосуглинистый, содержание гумуса — 3,9 %, подвижного фосфора и калия — соответственно 67,5 и 34,5 мг/100 г почвы, pH 5,0. Посев — по 500 всхожих семян на 1 м2 в три срока: 8, 16 и 25 мая (ГТК по срокам — соответственно 1,4; 1,9 и 2,7). Перед посевом в почву вносили нитроаммофоску — N40P40K40 и N120P120K120; контроль — без удобрений.
Каждый год опыты закладывали по методике ускоренного испытания генотипов (2). Задавали три уровня ГТК и три уровня NPK в полном факторном эксперименте. Таким образом, в один год вегетации (2000) было проведено 18 модельных испытаний шести сортов зерновых культур, различающихся соответственно 18 сочетаниями условий превегетации.
В период вегетации проводили фенологические наблюдения, измеряли длину (см) и ширину (мм), а также определяли площадь (см2) междоузлия, влагалища, листовой пластинки каждого метамера (от 1-го снизу к 9-му) в 10-15 повторностях. Для описания адаптационной способности растений в онтогенезе по методу В.М. Пасова (10) рассчитывали отношение длины предыдущего листа к длине последующего (Ln-1/Ln). После завершения вегетации выполняли полный структурный анализ биомассы растений и определяли урожайность зерна с 1 м2. В базу данных, помимо вышеупомянутых, включали метеорологические показатели накопления среднесуточных температур (°С) и суммы осадков (мм) за период вегетации и превегетации.
Данные обрабатывали статистически с использованием метода множественной регрессии.
Результаты. Полученная в эксперименте база данных по онтогенезу изученных сортов однолетних злаков с учетом условий среды в периоды превегетации и вегетации состояла из 100 столбцов и 15000 строк.
1. Влияние условий превегетации на урожайность сортов пшеницы и ячменя, рассчитанное методом множественной регрессии |
||||
Культура |
Сорт |
Фактор |
||
год формирования семян |
NPK |
ГТК |
||
Пшеница |
Ленинградка |
-0,21* |
0,08 |
-0,06 |
Крепыш |
0,19* |
0,14 |
-0,48* |
|
Иргина |
-0,24* |
0,06 |
-0,18* |
|
Ячмень |
Криничный |
-0,04 |
0,25* |
-0,07 |
Суздалец |
0,02 |
-0,02 |
-0,14 |
|
Эльф |
-0,09 |
0,06 |
-0,12 |
|
Доля случаев, % |
50 |
17 |
34 |
|
П р и м е ч а н и е. ГТК — гидротермический коэффициент. |
||||
Условия превегетации оказались более существенными для пшеницы, чем для ячменя (табл. 1). Год формирования семян влиял на урожайность чаще других факторов и поэтому был выбран в качестве критерия единственного различия семян.
|
Рис. 1. Урожайность зерна у сортов пшеницы (А) и ячменя (Б) в зависимости от года формирования семян и фона NPK. Варианты I и II — соответственно без внесения и с внесением удобрений. |
Наиболее выраженное действие условий превегетации на продуктивность растений пшеницы проявилось в контроле (без внесения удобрений) (в большей степени в случае сорта Крепыш по сравнению с двумя другими сортами) (рис. 1).
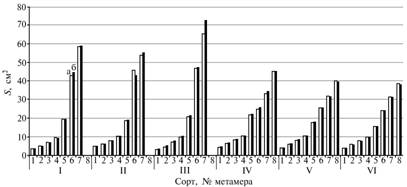
Несмотря на изменение массы растений, полученных из семян разных лет репродукции, не удалось обнаружить каких бы то ни было изменений в морфометрических характеристиках метамеров в связи с этим фактором. Незначительные флуктуации размеров междоузлия, влагалища листа и листовой пластинки были обнаружены в 5-7-м метамерах у всех сортов, но статистически значимые изменения по сортам, порядковому номеру метамера, части метамера, а также по характеристикам длины, ширины или площади метамера выявлены не были (исключение составил сорт Иргина, который реагировал на условия превегетации изменением роста последнего вегетативного метамера) (рис. 2). Это можно объяснить, во-первых, влиянием на продуктивность не только структуры главного, но и всех дополнительных побегов, во-вторых, изменением интенсивности физиологических процессов под влиянием учтенных условий превегетации без изменения морфометрических характеристик главного побега.
|
Рис. 2. Суммарная площадь поверхности частей метамера (междоузлие, влагалище, лист) у растений пшеницы и ячменя в зависимости от года формирования семян: I-VI — соответственно сорта пшеницы Ленинградка, Крепыш, Иргина и ячменя Криничный, Суздалец, Эльф; а и б — 1998 и 1999 годы; 1-8 — номер метамера. |
Характеристику приспособленности растений однолетних злаков к условиям среды выражали индексом адаптационной способности в онтогенезе: при оптимальных условиях каждая следующая листовая пластинка была длиннее предыдущей вплоть до развития последнего вегетативного метамера.
2. Уравнения регрессии отношения длины листовых пластинок (Ln-1/Ln) в зависимости от суммы накопленных температур (T, °С) для семян разных лет формирования у изученных сортов злаков |
||||
Сорт |
Год формирования семян |
Уравнение регрессии (T для потомственных растений) |
Т для |
Т1999 - Т1998 |
П ш е н и ц а |
||||
Ленинградка |
1998 |
Ln-1/Ln = 0,5962·exp(0,0009·T) |
575 |
6,3 |
1999 |
Ln-1/Ln = 0,6283·exp(0,0008·T) |
581 |
||
Крепыш |
1998 |
Ln-1/Ln = 0,6392·exp(0,0007·T) |
639 |
81,7 |
1999 |
Ln-1/Ln = 0,6488·exp(0,0006·T) |
721 |
||
Иргина |
1998 |
Ln-1/Ln = 0,6549·exp(0,0006·T) |
705 |
64,4 |
1999 |
Ln-1/Ln = 0,6805·exp(0,0005·T) |
770 |
||
Я ч м е н ь |
||||
Криничный |
1998 |
Ln-1/Ln = 0,4965·exp(0,0014·T) |
500 |
-12,8 |
1999 |
Ln-1/Ln = 0,5055·exp(0,0014·T) |
487 |
||
Суздалец |
1998 |
Ln-1/Ln = 0,5402·exp(0,0012·T) |
513 |
-12,7 |
1999 |
Ln-1/Ln = 0,5485·exp(0,0012·T) |
500 |
||
Эльф |
1998 |
Ln-1/Ln = 0,5328·exp(0,0013·T) |
484 |
16,5 |
1999 |
Ln-1/Ln = 0,5483·exp(0,0012·T) |
501 |
||
Используя этот подход, вычислили функции регрессии отношения длин листовых пластинок Ln-1/Ln в зависимости от суммы накопленных температур к моменту формирования листа исследуемого метамера (табл. 2). Как оказалось, влияние условий превегетации в период вегетации могло быть существенно на этапе формирования двух последних вегетативных метамеров у растений пшеницы. У растений из лучших семян продолжительность адаптивного роста предпоследнего и последнего метамеров увеличивалась на 0-3 сут (разность суммы температур 6,3-81,7 °C) в зависимости от сорта, что у сортов Крепыш и Иргина совпало с формированием большей массы зерна с колоса главного побега в контроле без удобрений. У растений ячменя период влияния года формирования семян на онтогенетическую адаптивность составлял не более 1 сут (разность суммы температур 12,7-16,5 °C) и приходился на формирование 6-го метамера, что практически не влияло на дальнейший рост растения и массу зерна с главного побега.
Описанные материнские фенотипические изменения не привели к значимому изменению биохимического состава семян в период вегетации (2), то есть условия вегетации нивелировали фенотипические различия по содержанию N, P и K в зрелых зерновках, вызванные действием факторов среды на этапе превегетации.
По нашим данным (8), у сорта Ленинградка (как у большинства сортов интенсивного типа — 12 из 13) в отличие от экстенсивного сорта Саратовская 29 посевные качества семян слабо зависели от экофизиологического последействия дозы удобрений, используемых на этапе превегетации. Напротив, по данным академика Е.И. Ермакова (3), при изучении урожайных свойств у четырех сортов пшеницы в регулируемой агроэкосистеме изменение температуры (12, 17, 22 °С) в период формирования семян снижало общую адаптивную способность и гомеостатичность растений сорта Ленинградка сильнее, чем сорта Саратовская 29. Следовательно, реакция сорта на условия превегетации зависит от генотипа и действующего внешнего фактора и выявляется в неодинаковой степени при использовании разных методов оценки экологического последействия. Кроме того, при подтверждении воздействия факторов превегетации на динамику развития растения, его продуктивность и другие характеристики необходимо контролировать генетическую однородность семян.
Существенное влияние условий превегетации на формирование 7-8-го метамеров по сравнению с 5-6-м у злаков может быть связано со скоростью роста междоузлий метамера. Другими авторами были описаны несортоспецифические особенности периодов скрытого и видимого роста вегетативных органов главного побега яровой пшеницы. Так, О.А. Евдокимова с соавт. (11) отмечали увеличение скорости роста верхних междоузлий относительно нижних. Н.А. Захарченко и В.А. Кумаков (12) сообщили о том, что в благоприятные годы период видимого роста листовой пластинки двух верхних вегетативных метамеров может увеличиваться на 2-3 сут по сравнению с острозасушливым годом. Аналогичным образом благоприятные условия превегетации продлевали период видимого роста листовых пластинок двух последних метамеров у пшеницы, что в дальнейшем приводило к повышению продуктивности растений. В нашем исследовании у растений ячменя изменение адаптивности из-за условий превегетации косвенно касалось только скрытого роста двух верхних вегетативных метамеров, а потому не оказывало существенного влияния на продуктивность растений.
Флуктуация длины междоузлия 5-7-го метамеров, выявленная у всех сортов пшеницы и ячменя, не зависела от условий превегетации, но была связана с порядковым номером метамера. При исследовании механической прочности стебля пшеницы также обнаружена разная длина междоузлий вегетативных метамеров (13), однако отмечено, что другие характеристики состояния метамера (максимальная сила среза или сгибания) мало зависели от порядкового номера метамера и были связаны с показателем относительной массы междоузлия на единицу длины. Для всех междоузлий, кроме верхнего, содержание гемицеллюлозы, целлюлозы и лигнина тоже мало зависело от порядкового номера метамера, но оказалось связано с величиной относительной массы междоузлия на единицу длины. Как известно, длина и прочность междоузлий вегетативных метамеров у злаков — достаточно устойчивый признак. В наших опытах условия превегетации (год формирования семян, NPK и ГТК) не влияли на длину междоузлий вегетативных метамеров.
Итак, на адаптивность растений яровой пшеницы наравне с условиями вегетации могут влиять условия превегетации. К таким условиям относятся год формирования семян и/или агроклиматический показатель влагообеспеченности (ГТК). Факторы превегетации значимы при формировании признаков продуктивности у пшеницы. Экофизиологическое последействие года формирования семян на адаптивность растений ячменя существенно ниже. Достоверного последействия влагообеспеченности материнских растений на урожайность сортов ячменя не обнаружено. Искажения результатов эколого-генетических экспериментов, связанные с условиями превегетации, меньше проявлялись у сортов ячменя, чем у сортов пшеницы. Размах этих искажений таков, что в контрольных опытах некоторые сорта давали разный результат по признакам продуктивности. Это может объясняться незначительным снижением/повышением адаптивности растений в период роста двух последних вегетативных метамеров в конце фазы выхода в трубку. Следовательно, как у пшеницы, так и у ячменя семена, полученные при применении традиционных агрохимических технологий и соблюдении допустимых пределов влажности почвы, могут быть использованы в эколого-генетических испытаниях. Если существует вероятность искажения результатов таких сортовых испытаний вследствие экологической разнокачественности семян, более информативно определение химического состава зерна и площади фотосинтезирующей поверхности главного побега. Эти признаки не подвержены материнскому фенотипическому эффекту даже при воздействии на растения лимитирующих факторов в период формирования семян. Адаптивность автогамных видов злаков обусловлена взаимодействием генотипа и среды, причем для некоторых генотипов среда превегетации может оказать дополнительное и существенное влияние на адаптационную способность в онтогенезе.
л и т е р а т у р а
ГНУ Агрофизический НИИ Россельхозакадемии, 195220 г. С.-Петербург, Гражданский просп., 14, а/я 63, |
Поступила в редакцию |
ADAPTABILITY IN CEREALS (Poaceae) IN CONNECTION WITH
PREVEGETATION AND VEGETATION CONDITIONS
N.A. Lykova
S u m m a r y
In the field experiments on three cultivars of summer wheat and on three cultivars of spring barley in different soil-climatic and ecological-geographic zones the author studied the effect of conditions of seed formation on their productive and adaptive properties in the period of vegetation of the offspring plants. The differences in reaction to seeds formation conditions were revealed as between Triticum aestivum L. and Hordeum vulgare L. species as between cultivars of the same species. It was shown that ontogenetic adaptability of wheat was changed by previous vegetation conditions in the moment of formation of the two last metamers (the seventh and eighth one from down). The control of hydrothermal conditions of maternal plants growth reduced significantly the risk of obstacles in ecological-genetic experiment with annual cereals.