СЕЛЬСКОХОЗЯЙСТВЕННАЯ БИОЛОГИЯ, 2008, № 1, с. 112-115
УДК 575.16
О РАЗРАБОТКАХ ТЕОРИИ ОНТОГЕНЕЗА РАСТЕНИЙ
И ЕЕ СВЯЗИ С БИОЛОГИЕЙ РАЗВИТИЯ
Ю.С. КОРЗИННИКОВ, А.А. ДОЛГОПОЛОВ
На основании обсуждения недостатков ряда теорий онтогенеза растений, предложенных за последние 50 лет, сделан вывод о неоправданности разработки какой-либо отдельной от биологии развития альтернативной теории онтогенеза.
Ключевые слова: биология развития, теория онтогенеза растений.
Key words: development biology, theory of plant ontogenesis.
Биология развития как общебиологическая дисциплина формировалась по двум направлениям — механика развития и генетика развития. Ее основные успехи были связаны с исследованиями, которые проводились на животных (Ч. Дарвин, Э. Геккель), послуживших первыми экспериментальными объектами (1, 2). Также предпринимались попытки продвинуть разработки в области биологии развития растений (3). Изучение механики развития растений проводили эмбриологи (В.А. Поддубная-Арнольди) и физиологи растений (Д.А. Сабинин, Н.П. Кренке, В.В. Скри-пчинский и др.). Что же касается генетического направления, которое является базисным, то отдельные частные исследования (4, 5) не могут служить тем фундаментом для разработки теории онтогенеза растений, каким был биогенетический закон (Ф. Мюллер и Э. Геккель, Ч. Дарвин) для биологии развития (онтогенеза) животных организмов. Итак, отправной точкой возникновения биологии развития можно считать открытие и разработку основного биогенетического закона. В этот период, вероятно, и сложился разделительный рубеж между отечественными теориями онтогенеза растений и биологией развития, в связи с чем стоит задача вернуть их разработку в русло биологии развития.
Отметим, что изучение онтогенеза растений — один из разделов биологии, в котором отечественные исследования не ориентировались на «лидерство» Запада, но при этом развивались в рамках сложившихся социальных особенностей и менталитета. Печально знаменитую теорию стадийного развития, относительно справедливую для ограниченного числа растений с низкотемпературным типом индукции генеративного этапа онтогенеза, неоправданно распространили на все царство растений как единственную и общепринятую: именно такой подход соответствовал на уровне социума условиям унитарного государства.
Накопленный объем знаний в этой области привел к размыванию теории стадийного развития и замене ее возрастной теорией, применимой для гораздо большего числа растений из разных систематических групп. Недостатком возрастной теории была постулированная тождественность онтогенеза растений с семенным и вегетативным способами возобновления, что противоречит генетической природе его регуляции (6). Возрастная теория прошла те же этапы становления и отмирания, что и теория стадийного развития: сначала экспериментальное изучение определенной группы растений, затем почти «законодательное» распространение сформулированных тезисов на все царство растений и, наконец, размывание под напором данных, полученных методами современной генетики.
В этот же период на базе изучения жизненного цикла многолетних травянистых растений развивалось так называемое популяционно-онтоге-нетическое направление (7-9). Как и обе упомянутые «унитарные» теории онтогенеза растений, популяционно-онтогенетическая претендует на некую всеобщность (10), то есть стремится пройти все этапы становления очередной унитарной теории. Закономерно возникает вопрос о целесообразности вычленения из биологии развития некой «сепаратной» теории онтогенеза растений.
Поскольку в геноме отсутствуют индивидуальные гены, ответственные за формирование той или иной структуры (увеличение числа клеток требует участия, как минимум, всего генома клеточного цикла), то сохранение в онтогенезе высших организмов формообразований, возникших сотни миллионов лет назад, указывает на устойчивое существование соответствующим образом организованных комплексов генов. Такие комплексы передаются или, по мнению Т.Г. Моргана (11), формируются в онтогенезе как целое. Первые многоклеточные организмы, как известно, были гаплоидными. У современных организмов гаплоидная фаза — гаметофит представляет собой одну из стадий жизненного цикла, а именно гаметогенез (образование половых клеток). Оплодотворение (образование спорофита — диплоидного поколения в виде покоящейся формы) появилось у растений в качестве адаптации к меняющимся условиям пересыхающих и перемерзающих пресноводных континентальных водоемов. То есть в сложных условиях наземного обитания стадии зиготы и спорофита возникли как альтернативный (запасной) вариант онтогенеза, который, однако, стал магистральным направлением морфологической эволюции высших форм организмов. Ароморфная эволюция спорофита, приведшая к его грандизации, оказалась возможной с возникновением регуляторного звена, определяющего временной ход развития, первым проявлением чего стало подавление перехода зиготы к редукционному делению без ограничения размножения диплоидных клеток (6). Такое подавление предполагает участие надиндивидуальных регуляторов генной активности, на роль которых могут претендовать гистоны.
У растений дифференциация исходной оси на органы происходит под индуктивным влиянием окружающих тканей и органов. Корень и лист можно считать модификациями оси (биполярной структуры) псилофитов (12). В онтогенезе цветковых особое место занимают семядоли. Они возникают на первом этапе дифференциации проэмбрио без участия формирующейся позже апикальной меристемы побега. По структуре и способу заложения семядоли цветковых растений идентичны первым листьям папоротников. Развитие корней у цветковых также остановилось на филогенетической ступени папоротников. С появлением у семенных растений стерильных листьев фертильные практически утратили фотосинтетические функции и превратились в микро- и мегаспорофиллы. У цветковых они становятся тычинкой и плодолистиком (5).
В онтогенезе высших растений мы не отмечаем появления фундаментальных жизненных функций, то есть их эволюция осуществлялась на основе структурных генов низших растений. При этом для возникновения зиготы как покоящейся стадии (половое размножение могло приобрести современное значение способа дифференциации потомства лишь после возникновения механизма рекомбинации) необходимы репрессоры, подавляющее функционирование генома в целом. Известно, что именно у зеленых водорослей появляется гистон, обладающий подобной способностью. Задержка редукционного деления, которая, как уже отмечалось, является первым эволюционным шагом спорофита, также предполагает участие надиндивидуальных регуляторов генной активности. Таким образом, геном исходной покоящейся зиготы стал первым «спорофитным» комплексом, а все остальные строились на его основе путем удвоения и эволюирования избыточного материала. Ароморфозы, появившись в филогенезе, остаются в онтогенезе эволюционно продвинутых групп. В онтогенезе цветковых зигота (арогенез водорослей) дает начало морфогенетически биполярному предзародышу (эволюционное приобретение псилофитов). Образующийся затем филлоризный зародыш («корень»—«гипокотиль–семядоли») является рекапитуляцией первичного тела папоротников, а стерильный побег — новообразованием. Тычинка и плодолистик представляют собой видоизмененные спорофиллы папоротников, семяпочка — упрощенный спорангий (напомним, что споры и гаметы возникли у водорослей).
Итак, мы перечислили этапы онтогенеза цветковых, естественно выделяемые при сравнительно-морфологическом изучении отделов ствола мегаэволюции покрытосеменных (рис.).
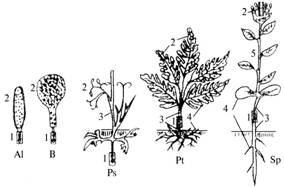
|
Оригинальная иллюстрация к ботаническому биогенетическому закону на примере появления новых морфологических структур спорофита, адаптированных к наземным условиям обитания: 1 — зигота, 2 — спорангий, 3 — морфогенетически биополярный кормус, 4 — филлоризный кормус, 5 — стерильный кормусный побег. Al — Algae, Br — Bryophyta, Ps — Psilopyta, Pt — Polypodiophyta, Sp — Spermatophyta. |
Поскольку гаметофит — наиболее древняя структура, его следует считать исходным элементом в онтогенезе цветковых растений (в норме — 8-ядерный зародышевый мешок и 2-ядерное пыльцевое зерно). Это правило осложняет морфологические исследования, но исключительно важно с точки зрения эволюции и функционирования генома.
В соответствии с изложенным онтогенез однолетних и многолетних растений как-либо принципиально не различается (имеются лишь особенности степени идиоадаптаций).
Многочисленные дискуссии и попытки применить основной биогенетический закон к царству растений (13) оказались безрезультатными, так как ботаники видели проявление биогенетического закона только в чередовании поколений (полового и бесполого) и не уделили внимания тому факту, что онтогенез, согласно биогенетическому закону, «начинается» с образованием зиготы, то есть касается только диплоидного организма или диплоидной фазы жизненного цикла. Возвращение разработок теории онтогенеза растений к этой отправной точке биологии развития (6, 14) осталось незамеченным. Гениальный W. Hofmeister (15) был ближе других к решению этой задачи, даже не углубляясь в детали основного биогенетического закона. Предложенная им на основе анализа морфологических признаков периодизация онтогенеза цветковых растений практически полностью совпадает с периодизацией, разработанной на методологической основе биогенетического закона (6, 16): зигота—биполярный предзародыш—филлоризный зародыш—кормус—спорофиллы—спорангии—споры—гаметы—зигота—…. Совпадение результатов, полученных разными методами, дает основание предполагать их объективность. Соглашаясь с критическим замечанием академика В.А. Драгавцева (17) об отсутствии завершенной теории онтогенеза растений, надеемся, что ее разработку ускорит именно возвращение исследований по этому научному направлению в область биологии развития.
Таким образом, теория онтогенеза растений может быть сформулирована только на основе и как часть общебиологической дисциплины —биологии развития.
Л И Т Е Р А Т У Р А
- Г и л ь б е р т С. Биология развития. М., 1993, т. 1, т. 2, т. 3.
- К о р о ч к и н Л.И. Введение в генетику развития. М., 1999.
- Биология развития растений. М., 1975: 229.
- Д р а г а в ц е в В.А., Л и т у н П.П., Ш к е л ь Н.М. и др. Модель эколого-генетического контроля. Докл. АН СССР, 1984, 274, 3: 720-723
- Д е м ь я н ч у к А.М. Расчет онтогенеза пшеницы. М., 2005.
- К о р з и н н и к о в Ю.С. К изучению генетико-морфологических вопросов онтогенеза растений. Изв. АН Тадж. ССР, отд. биол. наук, 1980, 4(81): 11-16.
- Р а б о т н о в Т.А. Жизненный цикл многолетних травянистых растений в луговых ценозах. Тр. БИН АН СССР, сер. 3 (Геоботаника). Л., 1950, вып. 6: 7-204.
- У р а н о в А.А. Возрастной спектр фитоценопопуляций как функция времени и энергетических волновых процессов. Биол. науки, 1975, 2: 7-34.
- Ценопопуляции растений (основные понятия). М., 1976.
- С м и р н о в а О.В., П а л ё н о в а М.М., К о м а р о в А.С. Онтогенез растений разных жизненных форм и особенности возрастной и пространственной структуры их популяций. Онтогенез, 2002, 33(1): 5-15.
- М о р г а н Т.Г. Развитие и наследственность. М., 1937.
- E m b e r g e r L. La valeur morphologique et l’oregone de la fleur. Ann. Biol. Linnee, 1950, 4(26).
- К о з о п о л я н с к и й Б.М. Основной биогенетический закон с ботанической точки зрения. Воронеж, 1937.
- К о р з и н н и к о в Ю.С. О принципе выделения этапов онтогенеза покрытосеменных растений. Докл. МОИП: Зоология и ботаника (1970-1-е полугодие 1971). М., 1972: 204-206.
- H o f m e i s t e r W. Allgemeine morpholgie des Gewächse. Leipzig, 1868.
- К о р з и н н и к о в Ю.С. Освоение генофонда и интродукция древесных плодовых растений на Западном Памире. Докт. дис. М., 1995.
- Д р а г а в ц е в В.А. Молекулярный, онтогенетический, популяционный и фитоценотический уровни эколого-генетической организации хозяйственно ценных признаков растений. С.-х. биол., 2006, 1: 115-123.
ФГОУ ВПО Иркутская государственная
сельскохозяйственная академия,
664038 Иркутская область, Иркутский район, п. Молодежный,
e-mail: rector@igsha.ru |
Поступила в редакцию16 ноября 2006 года
|
ABOUT WORKING OF THE PLANT ONTOGENESIS THEORY AND ITS RELATION WITH BIOLOGY OF DEVELOPMENT
Yu.S. Korzinnikov, A.A. Dolgopolov
S u m m a r y
On the basis of discussion of main disadvantages of different plant ontogenesis theories proposed during the latest 50 years the authors have made a conclusion regarding to unjustified another plant ontogenesis theory alternative to biology of development.