








УДК 579.869.1:56:575.829
К ФОРМИРОВАНИЮ БАКТЕРИЙ РОДА Listeria В ЭПОХУ ПОЗДНЕГО ПЛЕЙСТОЦЕНА: ФАКТЫ И ГИПОТЕЗЫ
В.И. ФЕРТИКОВ1, А.Н. ТИХОНОВ2, Е.М. ХРИПУНОВ3, И.Ю. ЕГОРОВА3
На основе обсуждения палеозоологических, палеонтологических и палеогеографических данных, характеризующих образование эколого-адаптивных механизмов у листерий, предложена гипотеза эволюции листерий и формирования у них свойств современных видов.
Ключевые слова: листерии, адаптация, изменение климата.
В природе существует огромное разнообразие микроорганизмов, из которых в настоящее время известно не более 5-7 %. Бактерии составляют их наиболее древнюю и многочисленную группу, возникшую примерно за 3-4 млрд лет до появления высших растений и животных. Первые формы микроорганизмов были исключительно свободноживущими (почвенные или водные сапрофиты). Возникновение более сложных форм жизни обусловило необходимость взаимодействия между ними и бактериями и привело к формированию новых взаимосвязей и взаимоотношений симбиотического и паразитического характера. Самый древний след подобной активности — поражение позвонка динозавра, найденного в палеозойских отложениях (300 млн лет назад). В отложениях мезозоя (100 млн лет назад) на позвоночнике крокодила обнаружены следы туберкулезного поражения (1). Бактерии заселяли поверхностные и внутренние экологические ниши других организмов, формируя новые экосистемы и играя определенную роль в естественном отборе животных и человека (2). Эволюция микроорганизмов, в частности их адаптация к многоклеточным организмам и паразитическому образу жизни, по мнению Л.П. Титова с соавт. (3), происходила в следующем направлении: свободноживущие сапрофиты, комменсалы и симбионты, внеклеточные, внутриклеточные и внутригеномные паразиты. В результате экологической эволюции на определенных территориях сформировались достаточно прочные (замкнутые или открытые) циклы циркуляций паразитических микроорганизмов в разнообразных популяциях животных. Можно предположить, что среди этих паразитирующих форм были и предшественники листерий, которые в настоящее время широко распространены в природе и как сапрофиты, и как патогены животных. Имеются сведения о выделении палочковидных форм бактерий (в том числе грампозитивных, к которым относятся листерии) из останков магаданского мамонтенка, погибшего 26 тыс. лет назад. Обнаруженные бактерии имели тот же размер и сходное строение поверхностных структур, что и некоторые современные бактерии — Escherichia coli, Bacillus subtilis, Pseudomonas aeruginosa и др. (4).
В «Определителе бактерий Берджи» (5) род Listeria отнесен к царству Бактерии, отделу Firmicutes, ко II классу Bacilli, порядку Bacillales, семейству Listeriaceae и включает 6 видов: L. monocytogenes, L. ivanovii, L. seeligeri, L. welshimeri, L. innocuaиL. grayi. На начальных этапах изучения листерий этот род включал лишь один вид — L. monocytogenes. Впоследствии были накоплены многочисленные данные о выделении листерий с различными биологическими свойствами, на основании чего в конце 1980-х годов сформировалась приведенная выше окончательная классификация рода Listeria.
Из перечисленных видов L. monocytogenes патогенна для животных и человека, L. ivanovii — только для животных. Этиологический статус L.se-eligeri до сих пор остается неясным, хотя в литературе имеются сведения о выделении этого микроорганизма из трупа человека, а также у больных с диагнозами ангина, менингит, инфекционный мононуклеоз, сепсис (6). Остальные три вида (L. welshimeri, L. innocua и L. grayi) непатогенны для животных и человека и широко распространены в природе. Существование в настоящее время паразитических и сапрофитических видов листерий, возможно, объясняется тем, что дивергенция (отделение) разных видов от общего предка проходила в разные периоды.
Ввиду того, что опасность для человека и животных представляет только вид L. monocytogenes, на нем проводилиосновные исследования биологических свойств листерий.
Э в о л ю ц и о н н о - а д а п т и в н ы е п р и з н а к и L. mono-cytogenes. Благодаря тому, что листерии обладают высокой экологической пластичностью и двойственной сапрофитно-паразитарной природой, они могут существовать в эндотермных и эктотермных организмах, растительных объектах, в почвенной и водной среде. В почве в зависимости от содержания гумуса, влажности и показателя рН происходит их активное размножение весной и осенью с перерывом летом. В хвойных лесах при почвенном рН 3,3-4,6 бактерии не размножаются, на полях без культивации остаются активными при рН 4,8-7,6. Листерии выделяли из растений, иксодовых клещей, 103 видов теплокровных и холоднокровных животных, обитающих во всех ландшафтных зонах, морских и речных системах, из ила и пыли. Один из эволюционных приспособительных механизмов, позволяющий листериям переживать неблагоприятные условия, — способность сохраняться в покоящихся цистах простейших (инфузорий), что служит проявлением симбиотических взаимоотношений по принципу «паразит—хозяин».
К эволюционно-адаптивным признакам листерий можно также отнести то, что при резком изменении средообразующих характеристик (температуры, влажности, состава питательных веществ и др.) они вырабатывают слизеподобную капсульную субстанцию, сохраняются и даже размножаются при пониженных температурах (группа психрофильных бактерий). В культурах некоторых штаммов после воздействия морской водой наблюдали 92,6 % капсулированных клеток. В зимний период L. monocytogenes остается жизнеспособной при температуре от -2 до -25 °С в течение 130 сут, во льду — до 2,5 лет; в морской и речной воде листерии размножаются при температуре от 1,5 до 44,0 °С. В талой пресной воде скорость роста бактериальной массы может увеличиваться в 2 раза. Кроме того, свидетельством высокой пластичности листерий служат данные о способности размножаться в присутствии высоких (до 20 %) концентраций солей, в то время как другие виды сапрофитных бактерий не выдерживают образующегося в морском льду повышенного уровня солености (7-12).
Адаптация L. monocytogenesи L. ivanoviiк макроорганизму привела к возникновению у животных и человека сапрозоонозной инфекции — листериоза, характеризующегося многообразием клинических форм и бессимптомным носительством (13). Листериоз человека входит в группу пищевых токсикоинфекций. По числу выявленных случаев он значительно уступает таким пищевым инфекциям, как сальмонеллез и кампилобактериоз, но превосходит их по летальности (20 %) и тяжести клинического течения.
В настоящее время листерии распространены во всем мире (Европа, Азия, Северная Америка, Африка) в различных климатических поясах и даже за полярным кругом, а листериоз как заболевание зарегистрирован в 56 странах. Первые сообщения о листериозе среди диких животных, в частности в популяциях песчанок (болезнь реки Tiger) в Южной Африке, описаны J.Y.Y. Pirie в 1927 году (14). В дальнейшем (работы, проведенные в большинстве случаев зарубежными исследователями) листерий обнаружили у енотовидной собаки, разновидностей мышей, косули, белохвостого оленя, лани, ламы, жирафа и др.
Учитывая отсутствие в доступной отечественной литературе данных о выделении листерий от диких животных на территории Российской Федерации, мы провели мониторинговые исследования фекалий диких парнокопытных в охотничьих угодьях некоторых областей Центрального региона России (табл. 1). Кроме того, при мониторинге распространения листерий у дикой фауны в водах Шошинского плеса Иваньковского водохранилища весной 2009 года мы обнаружили трупы четырех пятнистых оленей, утонувших осенью 2008 года. Методом полимеразной цепной реакции в тканях двух были выявлены фрагменты генома L. monocytogenes, а при бактериологическом исследовании одного из них изолировали культуру возбудителя листериоза.
| 1. Листерионосительство у видов диких парнокопытных животных в Центральном регионе России (январь—март 2009 года) | ||||
Место отбора проб |
Вид животного |
Число проб |
Выявлено листерионосителей |
|
всего |
листерионосительство, % |
|||
Калужская область, |
Кабан |
36 |
1 |
2,78 |
Пятнистый олень |
153 |
4 |
2,61 |
|
Марал |
5 |
0 |
0 |
|
Московская и Тверская области, Национальный парк «Завидово» |
Кабан |
39 |
1 |
2,56 |
Пятнистый олень |
97 |
1 |
1,03 |
|
Марал |
82 |
2 |
2,44 |
|
Лось |
9 |
0 |
0 |
|
Владимирская область |
Кабан |
25 |
3 |
12,00 |
Суздальское государственное опытно-охотничье хозяйство |
Пятнистый олень |
55 |
3 |
5,45 |
Лось |
2 |
0 |
0 |
|
Таким образом, изучение экологии листерий (ареал, взаимодействие в системах паразит—окружающая среда и паразит—макроорганизм)показывает, что их адаптационные механизмы позволяют размножаться в различных условиях и средах обитания, создавать стационарную природную очаговость болезни (15-17). Безусловно, формирование эколого-адаптивных механизмов, позволяющих L. monocytogenes переживать неблагоприятные воздействия, формировались в течение длительного исторического периода.
![]() У с л о в и я ф о р м и р о в а н и я э к о л о г о - а д а п т и в н ы х
У с л о в и я ф о р м и р о в а н и я э к о л о г о - а д а п т и в н ы х
м е х а н и з м о в у л и с т е р и й в э п о х у п о з д н е г о п л е й с т о
ц е н а (В а л д а й с к о е о л е д е н е н и е). В задачу настоящей статьи не входит изложение всех известных положений об истории четвертичного периода — мы будем рассматривать имеющиеся факты эволюции экологической адаптации листерий в связи с временными, климатическими и физико-географическими характеристиками Валдайского оледенения (рис. 1).
Образование Валдайского, Европейского (вюрм) и Висконсинского оледенения в Северной Америке вызвало понижение уровня Мирового океана на 130-150 м, вследствие чего Британские острова, Сахалин и Японские острова составляли одно целое с Евразийско-Американским материком. Зона тундростепей распространялась от Британской равнины через Среднюю Европу, Русскую равнину до Сахалина и Японии, на северо-востоке полностью захватывала Таймыр, Новосибирские острова, острова Франца-Ио-сифа, Аляску. Климат этого периода характеризовался малоснежной суровой зимой и холодным солнечным летом. Под воздействием климатических изменений и ледников возникли аридные приледниковые ландшафты — тундростепи, которые включали в себя элементы степной и тундровой растительности. Лессовые пылевые отложения обогащали почвы тундростепи, способствуя образованию мощной травянистой растительности под действием активной солнечной радиации, что в дальнейшем привело к формированию современных степей и черноземов. Сосна и береза первыми осваивали площади, освобождающиеся от ледников. Формирование хвойных и широколиственных лесов с участием березы, ольхи, лиственницы происходило только по долинам рек (19).
 |
Рис. 1. Валдайское оледенение (по Д.Д. Квасову, 1975) (18): 1 — максимальная стадия (17,0-15,0 тыс. лет назад), начальный период отступления ледников (16,0 тыс. лет назад); 2 — вепсовская стадия (15,5-14,5 тыс. лет назад); 3 — стадия салпаусселькая (11,8-10,2 тыс. лет назад); 4 — современная стадия. |
Состав поздней мамонтовой фауны по типу питания делился на две большие группы — травоядных и хи-щников (плотоядных), живущих за счет первых. Характерными для тундростепи были при-мерно 50 видов животных (20). Так, из позднего плейстоцена в го-лоцен перешли почти без заметных морфологических изменений из травоядных — благородный, пятнистый и северный олень, лось, косуля, кабан, сайгак, из хищников — волк, лисица, песец, росомаха, рысь, ряд видов грызунов.
В эпоху четвертичного периода (Рисс-Вюрм — потепление с исчезновением ледников на равнинах, длившееся примерно 50 тыс. лет) происходила смена целых фаунистических комплексов. Волжская (хазарская) степная фаза развития заменялась тундростепной (мамонтовой), которая длилась до начала голоцена. В этот период мамонты (мамонтовая фауна) в разное время заселяли всю Европу, Кавказ, северную половину Азии и часть Северной Америки, костные остатки этих животных находят в Атлантике в районе Нью-Йорка, на банках Северного моря.
Н.К. Верещагин (20-26) в своих работах обосновал тот факт, что изменения климата, рельефа, гидрографии, растительности, таяние ледников на всех континентах привели к гибели мамонтовой фауны. Известны ее прирусловые кладбища на крупных реках европейской части России, в Сибири и на Дальнем Востоке. Считается, что в основном гибель мамонтов происходила при переходах по непрочному льду рек, стариц, озер. В то же время палеонтологические данные о гибели парнокопытных и накоплении в кладбищах их останков немногочисленны.
В настоящее время Иваньковское водохранилище (1937 год) с Шошинским плесом на реке Волге можно рассматривать как модель «Тверского озера с Шошинским заливом» эпохи позднего плейстоцена (18, 27). Шошинский плес (8,2 тыс. га) с островами входит в состав национального парка. Острова в летне-осенний период используются парнокопытными (лось, марал, пятнистый олень, кабан) как места спаривания и выращивания молодняка. Поздней осенью по первому льду звери начинают с островов переходить в основной лесной массив к подкормочным площадкам. При фронтальных раннеосенних антициклонах с температурой от -10 до -15 °С и безветренной погодой на короткое время возникает «элемент аридности» (низкие температуры, солнечно и безветренно), Шошинский плес покрывается очень гладким льдом, на котором полностью отсутствует снежный покров. У парнокопытных при выходе на такой скользкий лед происходят разрывы паховых и тазобедренных связок и мышц, вывихи суставов, звери не могут встать и гибнут (по 10-15 особей за одну-две ночи). При отсутствии элемента аридности, то есть при теплой погоде и наличии снежного покрова на тонком льду, животные проваливаются в полыньи и также гибнут (в этом случае — единичные особи). Очевидно, так в «мамонтовых кладбищах» тысячелетиями накапливались останки копытных.
Можно предположить, что перемещение этих трупов весной по приледниковым водным системам к устьям рек, на побережье морей и океанов стало одним из путей распространения сапрофитной микрофлоры по морской литорали.
Проводя раскопки и обработку материалов «мамонтовых кладбищ», палеонтологи смогли установить примерную численность некоторых обитавших в четвертичный период видов животных. В частности, на основе архивных записей о продажах мамонтовой кости (бивни мамонта) в Восточной Сибири, а также данных раскопок и находок остатков мамонтовой фауны Н.К. Верещагин (24) оценил примерную численность мамонтов. По его данным, на протяжении последних 50 тыс. лет ледниковой эпохи на крайнем северо-востоке Сибири жило около 200 млн мамонтов. Каждый день мамонту требовалось примерно 300-400 кг растительной массы. В степях паслись сотенные косяки лошадей, ослов, тысячные стада бизонов, сайгаков, а ближе к ледникам передвигались бесконечные стада северных оленей. Всех этих животных, включая другие виды, обеспечивала кормами тундростепь с мощным травостоем из осок, злаков, полыней, пушицы, других растений и лишайников. Под их воздействием формировался гумусный слой степей. Хищникам в этот период также было достаточно пищи.
В Геохронологической лаборатории штата Аризона (США) изучили содержание 14С в костных остатках большого числа вымерших видов зверей из различных отложений и захоронений. Оказалось, что существовало несколько волн интенсивного вымирания мамонтовой фауны — 40 тыс., 13-8 тыс. и 4-0,4 тыс. лет назад. Они совпадали с периодами потепления климата, увеличением количества осадков и скорости таяния ледников, образованием приледниковых озер и рек, поднятием уровня Мирового океана на 50-80 м.
При таянии ледников и повышении уровня Мирового океана ряд приморских низменностей (включая Берингию с ее миллионами лошадей, бизонов и тысячами мамонтов) были затоплены. Сформировалась поверхность Земли в ее современном виде, отраженном на физико-географических картах. Сухопутная связь Евразии с Америкой через Берингию была прервана. Теплые зимы начали приносить многоснежье, гололеды и наледи, резко сокращалась возможность добычи корма растительноядными, что вело к перестройке всего фаунистического комплекса.
Потепление вызвало резкое изменение ландшафтов. Вместо продуктивных травянистых тундростепей формировались замшелые заозеренные тундры с обилием мхов и осок, болота с мириадами комаров, препятствовавших нагулу. В настоящее время эти жесточайшие условия тундры выдерживают лемминги, песцы и северные олени при условии миграции из лесотундры на побережье Ледовитого океана (25, 26, 28).
Для более полной фаунистической характеристики четвертичного периода необходимо хотя бы схематично рассмотреть воздействие ледников на образование внутренних водных систем (рек, озер, морей) и степень их засоленности. Установлено, что вследствие скопления в ледниках значительной части воды, участвующей в планетарном водообороте, снижался уровень Мирового океана. Кроме того, под влиянием оледенения происходили наиболее важные тектонические и изостатические движения позднечетвертичного периода — опускание и поднятие территорий, покрытых ледником.
Д.Д. Квасов (18) считает основными факторами развития озерных водных бассейнов Европы в позднечетвертичное время перестройку рельефа и гидрографической сети, вызванную непосредственно Валдайским оледенением; смену климата, который в период максимального оледенения, и в особенности в период позднего Сартанского оледенения (Вюрма), был сухим; изостатическое движение (в районе Балтийского и Белого морей, Ладожского и Онежского озер); изменение уровня Мирового океана (для Черного, Азовского, Балтийского, Белого морей) и рельефа дна Манычской долины, Керченского, Черноморских, Датских, Средне-Шведских проливов и горла Белого моря, а также водно-солевого режима Каспийского, Азовского, Черного и Балтийского морей.
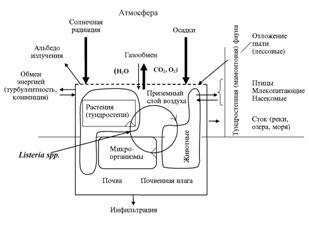 |
Рис. 2. Схема обмена энергии и вещества между биогеоценозом и окружающей средой (с участием Listeriaspp.) в позднем плейстоцене и голоцене (по материалам ссылок 19, 25, 29-31 с нашими дополнениями). |
Представленные данные о позднечетвертичном времени свидетельствуют, что на всех этапах на больших территориях изменялся кли-мат и характеристики физико-географической среды, растительного и животного мира, водных и водно-солевых бассейнов. Происходили процессы вымирания прежних и образования новых видов, их расселения, приспособления су-ществующих к новым условиям, формировались новые биогеоценозы, в которых активно участвовали и микроорганизмы (рис. 2).
Значительные изменения затронули растительные сообщества. Территория последнего Валдайского оледенения на протяжении всего четвертичного периода (вплоть до конца плейстоцена) претерпевала сложные геологические изменения, вследствие чего многократно сменялось действие различных по происхождению флорогенетических факторов — от субтропических до арктических и от континентальных до океанических. Здесь в течение всего позднего ледниковья происходили активные процессы формирования флористических комплексов Восточно-Европейской равнины (32). В то же время шли и обратные процессы приспособления уже сформировавшихся видов к новым условиям конца плейстоцена и начала голоцена. Так, многие группы мхов и сосудистых растений, найденных в кишечнике мамонтенка, погибшего в долине ручья Киргилях в Магаданской области (33), в настоящее время широко распространены в европейской и азиатской частях России (в частности, встречаются на территории Национального парка «Завидово» в Московской и Тверской областях) (табл. 2).
Таким образом, возможна следующая рабочая гипотеза, рассматривающая процесс формирования листерий в эпоху плейстоцена.
| 2. Зона распространения видов растений из кишечника мамонтенка, найденного в долине ручья Киргилях (Магаданская обл.), которые также встречаются на территории Национального парка «Завидово» (Московская и Тверская обл.) (цит. по 33; с нашими дополнениями) | ||
Вид |
Распространение |
|
общее |
в НП «Завидово» |
|
М х и |
||
Aulacomnium sp. |
ЕР, АР (распространен широко) |
Встречается часто |
Bryalesindet. |
ЕР, АР (распространен широко) |
Встречается часто |
Calliergon sp. |
ЕР, АР (распространен широко) |
Встречается часто |
Helodiumsp. |
ЕР, АР (встречается чаще на севере) |
Встречается редко |
Plagiomnium affine (Blandow ex Funck) |
ЕР, АР (распространен широко) |
Встречается часто |
Pleurozium sp. |
ЕР, АР (распространен широко) |
Встречается очень часто |
С о с у д и с т ы е р а с т е н и я |
||
Carex sp. |
ЕР, АР (распространен широко) |
Встречается часто |
Potentilla sp. |
ЕР, АР (распространен широко) |
Встречается часто |
Ranunculus sp. |
ЕР, АР (распространен широко) |
Встречается часто |
Ranunculus aff. flammula L. |
ЕР, АР (распространен широко) |
Встречается очень часто |
Rumex acetosella L. |
ЕР, АР (распространен широко) |
Встречается очень часто |
Selaginella aff. sibirica (Milde)Hieron. |
АР (Сибирь, Дальний Восток) |
Не встречается |
П р и м е ч а н и е. ЕР — Европейская Россия, АР — Азиатская Россия, НП — национальный парк. |
||
Вследствие аридности климата увеличился поток солнечной радиации, особенно в летний период, что ускоряло развитие травостоя в тундростепи и, как следствие, обусловило высокую численность млекопитающих мамонтовой фауны, а также образование в почве мощного гумусного горизонта. В то же время летняя солнечная радиация губительно действовала на листерий, которые в процессе эволюции приспособились к активному размножению весной и осенью (в периоды низких температур и уменьшения радиации), а также к существованию в растительном и гумусном слоях, защищающих их от радиации.
Условия длительных природных циклов четвертичного периода, огромная численность животных мамонтовой фауны способствовали формированию экологической пластичности и двойственной сапрофитно-па-разитарной природы некоторых видов Listeria spp., что через носительство обеспечило их переход из плейстоцена в голоцен.
Более активное размножение листерий в пресной воде, чем в соленой, служит свидетельством в пользу их изначальной «пресноводности» и вторичного приспособления к условиям засоленности, которое, возможно, происходило в период раннего голоцена (10,2-7,9 тыс. лет назад), когда менялась соленость водных бассейнов. Например, соленость Каспия резко возросла в результате прорыва соленых вод из Арало-Сарыкамышского бассейна, а в европейской части России — в связи со вторжением морской воды в Балтийскую котловину, поднятием уровня Мирового океана и последующим образованием соленых Черного и Азовского морей в их современных границах.
При образовании в четвертичный период кладбищ мамонтовой фауны приледниковые реки выносили трупы с сапрофитной микрофлорой в литоральную зону морей (океанов), где в дальнейшем они использовались эврибионтными организмами. Этот процесс происходил на всех континентах в ареале мамонтовой фауны, что подтверждается широким распространением Listeria spp. в биоте континентов и морей.
Итак, рассматривая становление экологической пластичности Listeriamonocytogenes, необходимо отметить, что она была сопряжена с процессами фауногенеза, происходившими под воздействием климатических и физико-географических пассажей четвертичного периода. Этим обусловлена целесообразность исследований фонотипических (прямых) механизмов экологической трансформации всего рода Listeria, связанных с факторами внешней среды и генетической структурой популяций.
Авторский коллектив выражает благодарность кандидату биологических наук, доценту Тверского государственного университета А.А. Нотову за консультации по мхам и сосудистым растениям.
Л И Т Е Р А Т У Р А
1. Ё л к и н И.И. Очерки теории эпидемиологии. М., 1960: 9-14.
2. П о з д е е в О.К. Медицинская микробиология. М., 2006.
3. Т и т о в Л.П., В о т я к о в В.И., К о ж е м я к и н А.К. и др. Эволюция микробов и ее медицинское значение. Здравоохранение, 2002, 8: 30-35.
4. Ш о ш а н и Дж., Г у д м а н М. и др. Клетки крови и белки мамонтенка, ультраструктура тканей и иммунодефицитное сравнение Mammuthus с ныне живущими Paenunguata. Магаданский мамонтенок. Л., 1981: 191-263.
5. Определитель бактерий Берджи. Т. 2. М., 1997: 574.
6. К р а с о в с к и й В.В. Листериозная инфекция на Украине. Кишечная картина заболевания. /http//www.ecologylife.ru/simpozium/listerioznaya-infektsiya-na-ukraine.html. 19.09.2008.
7. Б а к у л о в И.А. Листериоз сельскохозяйственных животных. М., 1967.
8. Х а у с м а н К. Протозоология. М., 1988.
9. М у х и н а Л.Б., Д м и т р и е в а Е.Ю. Организация контроля за распространением возбудителя листериоза на рыбоперерабатывающих предприятиях РФ. СПб, 2003: 7.
10. Б у з о л е в а Л.С., Т е р е х о в а В.Е. Выживаемость и адаптивная изменчивость штаммов Listeriamonocytogenesв морской и речной воде. Ветеринарная патология, 2004, 4: 31-35.
11. К о р е н б е р г Э.И. Преадаптивное происхождение возбудителей природноочаговых зоонозов. Усп. совр. биол., 2005, 125(2): 131-139.
12. П у ш к а р е в а В.И. Паразитизм в простейших как стратегия существования патогенных бактерий в почвах и водоемах. Усп. совр. биол., 2006, 126(4): 323-333.
13. Б а к у л о в И.А., В а с и л ь е в Д.А., К о л б а с о в Д.В. и др. Листерии и листериоз. Ульяновск, 2008.
14. P i r i e J.Y.Y. A new disease of wild rodents «Tiger River disease». Publication of the South African Institute for Medical Research, 1927, 3: 163-186.
15. Г р о м а ш е в с к и й Л.В. Общая эпидемиология. М., 1949.
16. С е р г и е в В.П., Т р е т ь я к о в А.Д. Инструкция о мероприятиях по профилактике и борьбе с листериозом животных. М., 1987: 61-67.
17. Б а к у л о в И.А., В а с и л ь е в Д.А. Листериоз как пищевая инфекция. Вопросы диагностики и профилактики. Ульяновск, 1991: 78.
18. К в а с о в Д.Д. Позднечетвертичная история озер и внутренних морей Восточной Европы. Л., 1975: 219-235.
19. В а л ь т е р Г. Растительность земного шара. М., 1974. Т. 2: 260-263.
20. В е р е щ а г и н Н.К. От ондатры до мамонта (Путь зоолога). СПб, 2002: 219-268.
21. В е р е щ а г и н Н.К. Великие «кладбища» животных в долинах рек Русской равнины. Природа, 1953, 12: 60-65.
22. В е р е щ а г и н Н.К. Охоты первобытного человек и вымирание плейстоценовых млекопитающих в СССР. Тр. зоол. ин-та АН СССР. Л., 1971. Т. 69: 200-231.
23. В е р е щ а г и н Н.К. О происхождении мамонтовых кладбищ. Природная обстановка и фауна прошлого (Киев), 1972, 6: 11-30.
24. В е р е щ а г и н Н.К. Гибель мамонтовой фауны в плейстоцене. Природа, 1977, 9: 90-95.
25. В е р е щ а г и н Н.К. Почему вымерли мамонты. Л., 1979.
26. В е р е щ а г и н Н.К., М а к е е в Ю.А. Происхождение и история северного оленя. В сб.: Северный олень в России 1982-2002 гг. М., 2003: 16-33.
27. Ф е р т и к о в В.И. Национальный парк «Завидово». М., 1998.
28. Ф е р т и к о в В.И., Т и х о н о в А.А., Н о в и к о в Б.Д. Современное состояние популяций и численность диких северных оленей в России. В сб.: Северный олень в России 1982-2002 гг. М., 2003: 56-73.
29. Ф л е р о в К.К. Млекопитающие и природная среда плейстоцена. Природа, 1977, 9: 50-56.
30. С у к а ч е в В.Н. Основы теории биогеоценологии. В юбилейном сб. АН СССР, посвященном Октябрьской революции. М.-Л., 1947: 30-46.
31. Д ы л и с С.В. Основы биогеоценоза. М., 1978: 6-123.
32. М и н я е в Н.А. История развития флоры северо-запада европейской части РСФСР с конца плейстоцена. Л., 1966: 38.
33. Н и к и т и н В.П. Остатки растений из кишечника мамонтенка. В сб.: Магаданский мамонтенок. Л., 1981: 242-289.
ON THE OCCASION OF FORMATION OF BACTERIA OF Listeria GENUS IN EPOCH OF LATE PLEISTOCENE: THE FACTS AND HYPOTHESIS
V.I. Fertikov1, A.N. Tikhonov2, E.M. Khripunov3, I.Yu. Egorova3
On the basis of discussion of the data of paleozoology, paleontology and paleogeography, characterized the appearance of ecologo-adaptive mechanisms in Listeria, the authors presented the hypothesis on listeria evolution and the formation in them the properties of modern species.
Key words: Listeria, adaptation, climate fluctuation.
1Национальный парк «Завидово», |
Поступила в редакцию |
Оформление электронного оттиска
![]()
![]()




