








УДК: 633.854.78:581.2:57.088.5
ПРИМЕНЕНИЕ RAPD-ПЦР МАРКЕРОВ ДЛЯ ДИФФЕРЕНЦИАЦИИ ФИЗИОЛОГИЧЕСКИХ РАС Plasmopara halstedii (Farl.) Berl. et de Toni, ПОРАЖАЮЩИХ ПОДСОЛНЕЧНИК В КРАСНОДАРСКОМ КРАЕ
С.З. ГУЧЕТЛЬ, Т.А. ЧЕЛЮСТНИКОВА, М.В. ИВЕБОР, Т.С. АНТОНОВА, Н.М. АРАСЛАНОВА, С.А. РАМАЗАНОВА
Методом полимеразной цепной реакции (ПЦР) с 22 RAPD-праймерами (RAPD — random amplified polymorphic DNA) проводили анализ молекулярно-генетического полиморфизма у возбудителя ложной мучнистой росы подсолнечника Plasmopara halstedii (Farl.) Berl. et de Toni для дифференциации пяти выделенных в Краснодарском крае рас патогена. Выявили, что четыре расы (330, 700, 710, 730) из пяти являются генетически близкими при четкой дифференциации расы 300.
Ключевые слова: ложная мучнистая роса, подсолнечник, Plasmopara halstedii, расы, ПЦР, RAPD, маркеры, полиморфизм, дифференциация.
Ложная мучнистая роса подсолнечника, вызываемая облигатным патогенным грибом Plasmopara halstedii (Farl.) Berl. et de Toniиз класса Oomycetes — одно из наиболее вредоносных заболеваний культуры во всем мире. По имеющимся данным, патоген характеризуется наличием более 30 патотипов. В последние годы с помощью стандартного набора линий-дифференциаторов подсолнечника, предложенного D. Tourvieille de Labro-uhe с соавт. (1), в отдельных регионах Северного Кавказа нами впервые идентифицировано 7 рас патогена. Однако такая дифференциация не всегда корректна и желательно иметь дополнительные маркеры для идентификации расовой принадлежности изолятов, используемых при тестировании устойчивости селекционного материала.
Из молекулярных методов анализа геномов (в том числе у грибов), позволяющих выявлять как межвидовую изменчивость, так и внутривидовые различия между патотипами, наиболее распространены модификации PCR-анализа (ПЦР-ДНК — полимеразная цепная реакция амплификации ДНК) (2, 3). Актуальным при изучении меж- и внутривидового разнообразия генома грибов остается RAPD-анализ ДНК (random amplified polymorphic DNA) (4, 5). Методом ПЦР с 30 RAPD-праймерами были обнаружены полиморфные фрагменты ДНК у 58 изолятов пяти патотипов P. halstedii, собранных воФранции (6). Это позволило подтвердить их дифференциацию на 5 рас. С помощью 21 RAPD-праймера была изучена взаимосвязь между расами 100, 300, 700, 330, 710, 703, 730, 770 из разных стран (7, 8). Выявлен слабый полиморфизм, который не зависит от расы и географического происхождения.
Возбудитель ложной мучнистой росы подсолнечника в России до недавнего времени оставался «белым пятном» как по расовому составу, так и по молекулярной характеристике генома. Ранее нами с использованием праймера на инвертированный повтор РТО были показаны различия между расами 330, 310, 710 и 730, выделенными на подсолнечнике в Краснодарском крае. По локусу РТО 450 у разных рас были выявлены контрастные аллельные состояния и внутрирасовый полиморфизм (9).
Цель настоящего исследования - анализ молекулярно-генетического полиморфизма и дифференциация пяти выделенных в Краснодарском крае рас P. halstedii с применением RAPD-ПЦР-маркеров.
Методика. Объектом исследований служили 30 изолятов пяти рас возбудителя ложной мучнистой росы — 300, 330, 700, 710 и 730 (соответственно 2, 12, 3, 6 и 7 изолятов), собранных с пораженных растений подсолнечника в разных районах Краснодарского края в 2004-2006 годах. Расовую принадлежность изолятов определяли в соответствии с номенклатурой, предложенной D. Tourvieille de Labrouhe с соавт. (1). Искусственное заражение линий-дифференциаторов и сортов подсолнечника осуществляли по общепринятой методике погружением корней проростков в инокулюм зооспор (10).
ДНК выделяли из конидиального спороношения P. halstedii по модифицированному методу М.Е. Zolan и P.J. Pukkila (11). Материал был собран с семядольных листьев проростков восприимчивого сорта подсолнечника ВНИИМК 8883, искусственно зараженных зооспорами каждого изолята отдельно.
Для RAPD-анализа использовали 16 десятичленных праймеров (Ope-ron Technologies, Inc., США) — OPM08, OPM20, OPJ15, OPB07, OPB17, OPC08, OPC15, OPD11, OPD18, OPE03, OPD20, OPF09, OPG02, OPG05, OPJ13, OPG06, позволивших продемонстрировать различия между пятью расами P. halstedii из разных областей Франции (6) и любезно предоставленных нам авторами. Кроме того, были испытаны шесть десятичленных праймеров, которые мы апробировали ранее на подсолнечнике (L11, Y14, P28, P38, P46, P53) (12).
ПЦР выполняли в реакционной смеси (25 мкл) следующего состава: 67 мМ Трис-HCl, рН 8,8; 16,6 мM сульфат аммония; 1,5-3,0 мM MgCl2; 0,01 % Tween 20; по 0,2 мM дезоксирибонуклеозидфосфатов; по 10 пМ праймеров; 10 нг матричной ДНК и 1 ед. рекомбинантной термостабильной ДНК-полимеразы («Госниигенетика», Россия). Амплификатор Терцик («АО ДНК-технология», Россия), режим — стандартный для RAPD-праймеров: начальная денатурация ДНК в течение 2 мин при 94 °С; 30 циклов с последовательной сменой температур: 60 с — 94 °С (денатурация), 60 с — 36 °С (отжиг), 120 с — 72 °С (элонгация); финальная элонгация 4 мин при 72 °С.
Электрофорез продуктов амплификации проводили в агарозном геле (1,5 % агароза, 1×ТАЕ-буфер) с использованием камеры для горизонтального электрофореза в течение 1,5-2,0 ч при I = 50 мА, U = 70-90 В. Реакционную смесь (10 мкл) наносили на гель вместе с красителем бромфеноловым синим. В качестве маркеров длин фрагментов ДНК использовали GeneRuler 1 kb DNA Ladder («Fermentas» UAB, Литва). Последующее окрашивание осуществляли бромистым этидием. Результаты электрофореза документировали при помощи трансиллюминатора и видеосистемы («АО ДНК-технология», Россия) с программным обеспечением Gel Imager 2. Поскольку при RAPD-ПЦР зачастую инициируется амплификация неспецифических фрагментов ДНК, реакции повторяли 2-3 раза, принимая за достоверные только воспроизводимые (в основном, мажорные) фракции. Различия между изолятами определялись по наличию или отсутствию полос на геле, соответствующих фрагментам ДНК определенной длины.
Меру сходства между расами рассчитывали с помощью коэффициента сходства Jacquard (13) по следующей формуле:
Jxy = nxy/(nx+ny),
где Jxy — коэффициент сходства Jacquard; nxy — число совпадающих по электрофоретической подвижности фрагментов у образцов X и Y; nx и ny — число амплифицированных фрагментов у образцов X и Y.
Результаты. Поскольку материал для экстракции ДНК P. halstedii был собран с растений подсолнечника сорта ВНИИМК 8883, на первых этапах работы для контроля отсутствия примеси ДНК хозяина в образцах дополнительно амплифицировали ДНК подсолнечника того же сорта. Воспроизводимые мажорные фрагменты ДНК гриба и подсолнечника всегда различались между собой по числу пар нуклеотидов, что свидетельствует о корректности сбора материала (рис., А).
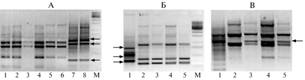 |
Электрофоретические спектры продуктов амплификации ДНК изолятов Plasmopara halstedi cразными RAPD-праймерами. А — с праймером L14 P. halstedii и подсолнечника: 1-6 — |
изоляты P. halstedii; 7 и 8 — сорт подсолнечника ВНИИМК 8883; стрелками отмечены воспроизводимые мажорные фракции длиной 550 п.н., 460 п.н. и 160 п.н. (сверху вниз), отличающие ДНК подсолнечника от ДНК гриба; Б — с праймером ОРJ13 рас P. halstedii: 1, 2, 3, 4 и 5 — соответственно расы 300, 330, 700, 710 и 730; стрелками отмечены фрагменты ДНК расы 300 длиной 710 п.н., 480 п.н. и 330 п.н. (сверху вниз), отличающие ее от остальных рас; В — с праймером ОРG06 рас P. halstedii: 1, 2, 3, 4 и 5 — соответственно расы 300, 330, 700, 710 и 730; стрелкой отмечена фракция 1125 п.н. М — маркер молекулярной массы ДНК (1 т.п.н.).
|
|
Из 22 использованных RAPD-праймеров один (Р38) не давал спектров амплифицированной ДНК. Остальные инициировали амплификацию, в общей сложности, 92 фрагментов со средней частотой 4,4 фрагмента на праймер. Восемь праймеров (L14, Y11, P28, Р46, P53, OPM08, OPM20, OPJ15) не выявили полиморфизма у изолятов, с тринадцатью (OPB07, OPB17, OPC08, OPC15, OPD11, OPD18, OPE03, OPD20, OPF09, OPG02, OPG05, OPG06 и OPJ13) было обнаружено отличие изолята Д8-5-05расы 300 от остальных четырех анализируемых рас, что проявлялось либо в наличии фракций амплифицированной ДНК, отсутствующих у других рас, либо в отсутствии фракций, типичных для последних (см. рис., Б). Всего эти тринадцать праймеров продуцировали 76 фрагментов (от 1 до 12 на праймер), выявив 62 полиморфных локуса.
Только с одним праймером (ОРG06) был показан внутрирасовый полиморфизм по наличию/отсутствию фракции фрагмента ДНК длиной 1125 п.н. (обозначена как ОРG061125) (см. рис., В). Два изолята расы 300, собранные в разных районах Краснодарского края, обладали по этому признаку разным генотипом. Из шести изолятов расы 330 два имели данный фрагмент ДНК (оба в отличие от других были собраны на полях ВНИИМК) (табл.). Внутрирасовую изменчивость по этому признаку проявила также раса 730: в спектре фрагментов амплифицированной ДНК фракция ОРG061125 была обнаружена у изолята Д2-17-05; у остальных она отсутствовала. Не было выявлено внутрирасовой изменчивости у изолятов расы 710: у всех изученных (см. табл.) указанная фракция присутствовала в спектре, что свидетельствует о некой молекулярной стабильности этого локуса у указанной расы по сравнению с остальными.
Следует отметить, что при изучении внутривидовой вариабельности французских изолятов рас 100, 300, 700, 703 и 710 P. halstedii не показана внутрирасовая изменчивость (6). В то же время внутрирасовые различия были обнаружены у одноименных рас, происходящих из разных стран. У изолятов одного географического происхождения эти различия иногда так существенны, что образцы относятся к разным кластерам дендрограммы, построенной на основе оценки генетических дистанций между ними (8). Наличие внутрирасовой изменчивости может свидетельствовать о возможном гибридном происхождении рас. Подобные полиморфные RAPD-локусы, по-видимому, не сцеплены с генами вирулентности гриба — в этом случае в популяции могут сосуществовать особи с разным аллельным состоянием указанных локусов.
Характеристика изолятов Plasmopara halstedii, проанализированных по локусу ОРG061125 с применением RAPD-ПЦР-маркеров |
|||
Раса |
Название |
ОРG061125 |
Место сбора изолята |
300 |
Д8-5-05 |
+ |
г. Краснодар, поля ВНИИМК |
300 |
Л-1-05 |
- |
Лабинский р-н |
330 |
ВК-8-05 |
- |
г. Краснодар, поля ВНИИМК |
330 |
ВК-2-06 |
+ |
|
330 |
А-131-05 |
- |
Каневской р-н, станица |
330 |
Ф1-2-05 |
- |
Каневской р-н, фермерское поле |
330 |
В-61-05 |
+ |
Выселковский р-н, поле 1 |
330 |
В-26-05 |
- |
|
700 |
Л-5-05 |
+ |
Лабинский р-н |
700 |
В-П2-05 |
+ |
Выселковский р-н, поле 2 |
710 |
В-85-13 |
+ |
То же |
710 |
В-89-05 |
+ |
|
710 |
ВК-13-05 |
+ |
г. Краснодар, поля ВНИИМК |
710 |
Гл-2-05 |
+ |
Неизвестно |
730 |
Гл-3-05 |
- |
Неизвестно |
730 |
В-46-05 |
- |
Станица Выселки |
730 |
Д2-16-05 |
- |
г. Краснодар, поля ВНИИМК |
730 |
Д-17-05 |
+ |
|
730 |
ВК-2-05 |
- |
|
П р и м е ч а н и е. «+» — наличие, «-» — отсутствие у образца амплифицированного фрагмента; ВНИИМК — Всероссийский НИИ масличных культур. |
|||
В наших опытах из пяти собранных в Краснодарском крае и проанализированных по 62 полиморфным локусам рас P. halstedii четко дифференцировалась только раса 300. Мы определили меру сходства между расой 300 и остальными расами с помощью коэффициента сходства Jxy (этот метод расчета предпочтительно использовать при обработке данных RAPD) (14). Как известно, коэффициент может принимать значения от 0 для совершенно различных генотипов до 1 для идентичных генотипов. Мера сходства между расами 330, 700, 710 и 730 была близка к 1 (что объединило их в одну группу), между расой 300 и другими расами в нашем эксперименте определялась достаточно малой величиной — 0,29. Соответственно генетическая дистанция (Dxy) между расой 300 и остальными расами, рассчитываемая по формуле Dxy = 1 - Jxy, составляла 0,71. Для сравнения: уровень сходства между расами 300 и 710 из Франции — 0,88 при наибольшем значении для «старых» рас 100 и 300 — 0,99 (6). В настоящее время раса 100, которая наименее вирулентна, встречается в Краснодарском крае спорадически, что не позволило охарактеризовать ее генотип и степень близости с другими патотипами.
Отличие расы 300 от остальных вполне закономерно. Хотя расовый состав патогена не отслеживался в течение долгого времени, по данным исследований, имеющимся на начало 80-х годов ХХ столетия, можно предположить, что раса 300 появилась в Краснодарском крае вслед за расой 100 (10). Напомним, что по старой номенклатуре это раса 2, известная как Red River Valley и обнаруженная впервые в долине одноименной реки в Северной Америке и Канаде (15). Известно, что в Европе была распространена только раса 1 (или 100 по новой номенклатуре). Происхождением расы 300 с другого континента можно объяснить быстрое выявление ее отличия от остальных по изученным локусам. Наши данные согласуются с исследованиями других авторов (7), сгруппировавших в отдельные кластеры европейские и американские изоляты гриба.
При изучении рас P. halstedii, происходящих с четырех континентов, обнаружено (8), что все изоляты расы 100 группируются в отдельный кластер, то есть генетически отдалены от других. Авторы предполагают, что все «новые» расы происходят не от расы 100, а от другой — возможно, от расы 300. Как отмечалось выше, у нас не было возможности изучить расу 100, происходящую из Краснодарского края, однако очевидно, что «новые» расы (330, 700, 710 и 730) генетически отдалены от «старой» расы 300 и, вероятно, не имеют с ней общего происхождения. Ограниченное число проанализированных изолятов указанной расы не позволяет считать этот факт неоспоримым: так же, как раса 100, в настоящее время раса 300 встречается в Краснодарском крае очень редко и дает скудное спороношение, что затрудняет ее поддержание в коллекции. Большая степень сходства между расами 330, 700, 710 и 730 свидетельствует об их общем происхождении, то есть может иметь место «эффект основателя» — происхождение от небольшого числа генотипов. И хотя в наших экспериментах у этих рас не показаны межрасовые различия по изученным локусам ДНК, не исключена возможность их дифференциации по другим локусам, находящимся в одной группе сцепления со специфическими генами вирулентности.
Таким образом, с использованием RAPD-ПЦР-маркеров показано, что из пяти рас Plasmopara halstedii, собранных на подсолнечнике в Краснодарском крае, у четырех (330, 700, 710 и 730) уровень сходства близок к 1, что может свидетельствовать об общности их происхождения. По 21 RAPD-праймеру четко дифференцируется раса 300: уровень сходства между этой расой и остальными равен 0,29. По локусу ОРG061125 обнаружена внутрирасовая изменчивость у изолятов рас 300, 330, 700 и 730 и мономорфность — у изолятов расы 710.
Л И Т Е Р А Т У Р А
1. T o u r v i e i l l e d e L a b r o u h e D., G u l y a T.J., R a s h i d Y.K. e.a. New nomenclature of race of Plasmopara halstedii (sunflower downy mildew). Proc. 15th Internat. sunflower conf. Toulouse, France, 2000, 2: 161-166.
2. S i v a r a m a k r i s h n a n S., K a n n a n S e e t h a, S i n g S.D. Genetic variability of Fusarium wilt pathogen isolates of chickpea (Cicer arientinum L.) assessed by molecular markers. Micopatologia, 2002, 155(3): 171-178.
3. G e r m a i n H., L a f l a m m e G., B e r n i e r L. e.a. DNA polymorphism and molecular diagnosis in Inonotus spp. Can. J. Plant. Pathol., 2002, 24(2): 194-199.
4. Ш н ы р е в а А.В., Ш т а е р О.В. Дифференциация близкородственных видов Pleurotuspulmonaris и P. osteratus с помощью скрещиваний и молекулярных маркеров. Генетика, 2006, 42(5): 667-674.
5. M a r i n M., C a s t r o B., G a i t a n A. e.a. Relationships of Ceratocystis fimbriata isolates from Colombian coffee growing regions based on molecular data and pathogenecity. J. Phytopatol., 2003, 151(7-8): 395-405.
6. R o e c k e l-D r e v e t P., C o e l n o V., T o u r v i e i l l e J. e.a. Lack of genetic variability in French identified races of Plasmopara halstedii, the cause of downy mildew in sunflower Helianthus annuus. Can. J. Microbiol., 1997, 43: 260-263.
7. T o u r v i e i l l e J., M i l l o n J., R o e c k e l-D r e v e t P. e.a. Molecular variability of Plasmopara halstedii. Proc. 15th Internat. sunflower conf. Toulouse, France, 2000: 67-72.
8. R o e c k e l-D r e v e t P., T o u r v i e i l l e J., G u l y a T.J. e.a. Molecular variability of sunflower downy mildew, Plasmopara halstedii from different continents. Can. J. Microbiol., 2003, 49(8): 492-502.
9. Г у ч е т л ь С.З., А н т о н о в а Т.С., А р а с л а н о в а Н.М. и др. Первый опыт выявления молекулярно-генетических различий у патотипов возбудителя ложной мучнистой росы, поражающей подсолнечник на Северном Кавказе. Тр. II Всерос. съезда по защите растений. СПб, 2005, 1: 158-160.
10. А н т о н о в а Т.С., А р а с л а н о в а Н.М., Г о л о в и н А.В. и др. К вопросу о расовой принадлежности возбудителя ложной мучнистой росы на подсолнечнике на Северном Кавказе. Науч.-техн. бюл. ВНИИМК, 2000, 123: 16-20.
11. Z o l a n M.E., P u k k i l a P.J. Inheritance of DNA methylation in Corpinus cinereous. Mol. Cell Biol., 1986, 6(1): 195-200.
12. Г у ч е т л ь С.З., Ч е л ю с т н и к о в а Т.А., Р а м а з а н о в а С.А. и др. Молекулярно-генетическая характеристика инбредных линий подсолнечника по изоферментным маркерам и ДНК-профилям. Науч.-техн. бюл. ВНИИМК, 2004, 2(131): 42-46.
13. S n e a t h P.H., S o c a l R.R. Numerical taxonomy: the principles and practice of numerical classification. San Francisco, 1973.
14. L i n k W., D i x k e n s C., S i n g h M. e.a. Genetic diversity in European and Mediterranean faba bean plasma reveal by RAPD markers. Theor. Appl. Genet., 1995, 90: 27-32.
15. Z i m m e r D., K i n m a n M.L. Downy mildew resistance in cultivated sunflower and its inheritance. Crop Sci., 1972, 12: 749-751.
APPLICATION OF RAPD-PCR MARKERS FOR DIFFERENTIATION OF PHYSIOLOGICAL RACES IN Plasmopara halstedii (Farl.) Berl. et de Toni INFECTING SUNFLOWER IN KRASNODARSKII REGION
S.Z. Guchetl’, T.A. Chelyustnikova, M.V. Ivebor, T.S. Antonova, N.M. Araslanova, S.A. Ramazanova
By the method of DNA polymerase chain reaction (PCR) with 22 RAPD-primers (RAPD — random amplified polymorphic DNA) the authors analyzed the molecular-genetic polymorphism in Plasmopara halstedii (Farl.) Berl. et de Toni, an agent of mildew of sunflower, for differentiation of five pathogen races isolated in Krasnodarskii region. It was revealed, that four races (330, 700, 710, 730) from five are similar genetically with sharply defined race 300.
Key words: downy mildew, sunflower, Plasmopara halstedii, races, PCR, RAPD, mark-ers, polymorphism, differentiation.
ГНУ Всероссийский НИИ масличных культур |
Поступила в редакцию |
Оформление электронного оттиска
![]()
![]()




