








УДК 636.52/58:575.174
ГЕНЕТИЧЕСКАЯ ГЕТЕРОГЕННОСТЬ И ДИВЕРГЕНЦИЯ ПОРОД КУР, ИМЕЮЩИХ КОРНИ БОЙЦОВОЙ ПТИЦЫ
А.Ф. ЯКОВЛЕВ, В.И. ТЫЩЕНКО, Н.В. ДЕМЕНТЬЕВА, О.В. МИТРОФАНОВА, В.П. ТЕРЛЕЦКИЙ, А.Б. ВАХРАМЕЕВ
Методом фингерпринтинга ДНК определяли внутрипородные и межпородные коэффициенты генетического сходства, генетические расстояния и степень гетерозиготности у кур пород юрловская голосистая, орловская ситцевая, узбекская бойцовая и московская бойцовая, имеющих генетические корни бойцовых кур. Наиболее генетически отдаленной от трех изучаемых пород оказалась орловская ситцевая. Степень гетерозиготности у кур этой породы также была наиболее низкой. У изученных пород выявлены маркерные фрагменты ДНК (или их отсутствие). Обсуждаются проблемы генетического взаимоотношения четырех изучаемых пород в связи с их принадлежностью к бойцовому типу.
Ключевые слова: полиморфизм ДНК, ДНК-фингерпринтинг, породы кур, генетические расстояния, гетерозиготность.
Молекулярные маркеры, основанные на полиморфизме ДНК и генов, в настоящее время приобретают ключевую роль в генетике сельскохозяйственных животных. Имеется достаточно широкий выбор методических подходов к изучению различных полиморфных систем ДНК в зависимости от целей исследования. Наибольший вклад в оценку генома животных вносят методы полимеразной цепной реакции (ПЦР), капиллярного электрофореза и ДНК-фингерпринтинга. Последний выявляет полиморфизм микросателлитных фрагментов ДНК с различной длиной (или молекулярной массой). Если в качестве основной цели стоит оценка племенного поголовья, то необходимо учитывать как простоту выполняемого теста, так и его стоимость.
Создание высокопродуктивных промышленных пород приводит к постепенному вытеснению исходного генофонда старых, местных пород —носителей ценного генетического материала, который часто остается невостребованным. Изменение условий содержания животных, конъюнктурных требований рынка, появление каких-либо других факторов в будущем потребует привлечения генетических ресурсов от этих старых пород. К сожалению, поголовье таких животных неуклонно снижается и в ряде случаев само существование пород ставится под сомнение. При сохранении пород следует поддерживать высокую численность популяции, что экономически не выгодно. И в этом случае большую помощь может оказать контроль генетического разнообразия и дивергенции популяций при помощи ДНК-технологий.
Полиморфные ДНК-маркеры были использованы для идентификации пород кур различного происхождения (1, 2), разработаны приемы оценки генетического разнообразия в породах и линиях кур (3, 4). Сделана попытка связать полиморфные системы одиночных нуклеотидов у линий кур с хозяйственными признаками (5). С помощью анализа полиморфизма микросателлитной ДНК достаточно точно измерена степень дивергенции между современными и древними породами кур (6). Технология фингерпринтинга ДНК как способ контроля за генофондными популяциями кур была использована нами при анализе промышленного стада птицы (7). Этот метод дает возможность оценить генетическое разнообразие пород, линий и популяций, выявить межпородный и внутрипородный полиморфизм для оценки генетического взаимоотношения пород.
Настоящая работа посвящена выявлению внутрипородной и межпородной генетической гетерогенности и дивергенции у птицы, имеющей корни бойцовых кур.
Методика. Исследование проводили на курах генофондных популяций четырех пород: юрловская голосистая, орловская ситцевая, узбекская бойцовая, московская бойцовая (Экспериментальное хозяйство ГНУ ВНИИГРЖ, Ленинградская обл.). Для оценки генетического разнообразия применяли метод ДНК-фингерпринтинга (3), который включал выделение ДНК, расщепление ДНК рестриктазами, электрофорез фрагментов в агарозном геле («Sigma», США), их перенос с агарозного геля на фильтр, предгибридизацию и гибридизацию ДНК, выявление дезоксигенина на фильтрах, анализ распределения гибридизационных фрагментов на фильтрах («Amersham», Великобритания). Использовали зонд (GTG)5 (то есть последовательность G-T-G повторяется 5-кратно).
Популяционно-генетические характеристики — число полос на дорожку, вероятность встречаемости двух одинаковых генотипов (Р), коэффициенты генетического сходства внутри групп (BS1) и между группами (BS2), генетические расстояния (D), число локусов, число аллелей на один локус и степень гетерозиготности — рассчитывали с помощью компьютерной программы Gelstats (3). При определении размера выборки исходили из рекомендаций, разработанных для проведения исследований методом фингерпринтинга ДНК (8), согласно которым достоверные данные получают при величине выборки 10-15 гол.
Результаты.Краткая характеристика изучаемых пород свидетельствует, что все они имеют генетические корни бойцовых кур (табл. 1).
| 1. Характеристика генофондных пород кур (Экспериментальное хозяйство ГНУ ВНИИГРЖ, Ленинградская обл.) | |||||
Порода |
Порода, фактически/пред-положительно участвовавшая в выведении |
Тип |
Показатель продуктивности |
||
яйценоскость, шт/год |
живая |
масса |
|||
Юрловская |
Брама, бойцовые |
Мясо-яичный |
160-200 |
2,6-3,0 3,0-3,5 |
58-62 |
Орловская |
Бойцовые малайские |
Декоративно-бойцовый |
140-160 |
1,6-2,7 2,4-3,5 |
51-52 |
Узбекская |
Местные куры из |
Бойцовый |
110-130 |
2,4-2,8 2,8-3,2 |
53-57 |
Московская |
Бельгийские и малайские бойцовые куры |
Бойцовый |
150-160 |
2,2-3,0 2,7-3,5 |
55-57 |
Полученные гибридизационные спектры свидетельствовали о высоком генетическом разнообразии как между индивидами внутри породных групп, так и между породами. Межпородный полиморфизм проявлялся в виде различий по числу полос, их распределению, интенсивности и ширине отдельных полос. Наиболее высокий индивидуальный полиморфизм отмечали в области фрагментов ДНК длиной от 7 до 20 т.п.н. Как у индивидов, так и у пород в зоне фильтров, соответствующей размеру фрагментов 2-7 т.п.н., имелись блоки сходных полос. В целом отмечалась тенденция к снижению частоты встречаемости полиморфных полос по мере уменьшения длины фрагмента.
Вероятность встречаемости двух особей с одинаковым набором фрагментов ДНК (Р) составляла один случай на несколько миллиардов голов, что свидетельствует о высокой точности метода (табл. 2). Была выявлена удаленность орловской ситцевой породы от трех других.
| 2. Внутрипородные и межпородные коэффициенты сходства (BS) и генетические расстояния (D) у кур из генофондных популяций изучаемых пород, имеющих генетические корни бойцовой птицы (по данным ДНК-фингер-принтинга; Экспериментальное хозяйство ГНУ ВНИИГРЖ, Ленинградская обл.) | ||||||
Сравниваемая |
n |
Число полос на |
P |
BS1 |
BS2 |
D |
Юрловская голосистая/ |
10 |
37,2±2,0 |
1,8x10-18 |
0,33 |
0,28 |
0,100 |
Юрловская голосистая/ Узбекская бойцовая |
10 |
37,2±2,0 |
1,8x10-18 |
0,33 |
0,28 |
0,070 |
Юрловская голосистая/ Московская бойцовая |
10 |
37,2±2,0 |
1,8x10-18 |
0,33 |
0,29 |
0,050 |
Орловская ситцевая/ |
10 |
30,1±1,1 |
5,7x10-12 |
0,42 |
0,25 |
0,140 |
Орловская ситцевая/ |
10 |
30,1±1,1 |
5,7x10-12 |
0,42 |
0,29 |
0,090 |
Узбекская бойцовая/ |
11 |
33,4±1,5 |
1,7x10-15 |
0,36 |
0,29 |
0,060 |
П р и м е ч а н и е. P — вероятность встречаемости двух особей с одинаковым набором фрагментов ДНК, BS1 и BS2 — коэффициенты сходства соответственно внутри и между породами. |
||||||
Группы кур пород юрловская голосистая и московская бойцовая характеризовались высоким генетическим сходством и, соответственно, низкими значениями генетического расстояния (D = 0,050). Примерно одинаковые небольшие генетические расстояния были отмечены между узбекской бойцовой и двумя породами — московской бойцовой (D = 0,060) и юрловской голосистой (D = 0,070). Однако при значительном генетическом сходстве между породами имелись различия по частоте некоторых фрагментов
| 3. Частота некоторых фрагментов ДНК у кур из генофондных популяций изучаемых пород (по данным ДНК-фингерпринтинга; Экспериментальное хозяйство ГНУ ВНИИГРЖ, Ленинградская обл.) | ||||
№ фрагмента ДНК |
Юрловская |
Орловская ситцевая |
Узбекская |
Московская |
13 |
0,80 |
0,10 |
0 |
0,27 |
52 |
0,20 |
0,60 |
0,36 |
0,91 |
106 |
0,40 |
0,90 |
0,09 |
0,64 |
114 |
0,30 |
0,80 |
0,09 |
0,36 |
120 |
0,30 |
1 |
0 |
0,27 |
123 |
0,50 |
0 |
0,73 |
0,64 |
ДНК (табл. 3). Так, несмотря на обмен генетическим материалом между юр-
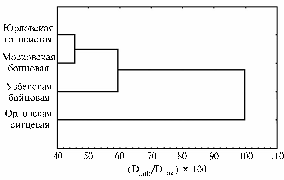 |
Дендрограмма генофондных популяций кур изучаемых пород, имеющих генетические корни бойцовой птицы (по данным ДНК-фингерпринтинга; Экспериментальное хозяйство ГНУ ВНИИГРЖ, Ленинградская обл.). |
ловской голосистой и бойцовыми породами, каждая характеризовалась отличительными особенностями, свойственными для изучаемой популяции птицы. Например, у кур породы юрловская голосистая выявили фрагмент № 13, который встречается у 80 % особей и может служить маркером породы. В по-пуляции кур узбекской бой-цовой породы он не встречался. Для орловских кур были типичны фрагменты № 106 и № 120, а фрагмент № 123 у этой породы не наблюдали. Генетические взаимоотношения между изучаемыми породами, в частности некоторую обособленность узбекской бойцовой и особенно орловской ситцевой пород, иллюстрирует дендрограмма (рис.).
Расчет показателей гетерозиготности (табл. 4) продемонстрировал, что в выборке кур орловской ситцевой породы генетическая гетерогенность наиболее низкая по сравнению с тремя другими изучаемыми породами, имевшими примерно одинаковые показатели гетерозиготности. Степень гетерозиготности, в свою очередь, отражает характер разведения птицы в изучаемой популяции.
| 4. Показатели гетерозиготности у кур из генофондных популяций изучаемых пород, имеющих генетические корни бойцовой птицы (по данным ДНК-фингерпринтинга; Экспериментальное хозяйство ГНУ ВНИИГРЖ, Ленинградская обл.) | |||||||
Порода |
n |
Число |
H1 |
H2 |
H3 |
||
локусов |
аллелей |
полиморфных |
|||||
Юрловская голосистая |
10 |
21,19 |
5,66 |
1,00 |
0,76 |
0,86 |
0,81 |
Орловская ситцевая |
10 |
18,21 |
5,11 |
0,95 |
0,65 |
0,75 |
0,71 |
Узбекская бойцовая |
11 |
19,23 |
5,72 |
1,00 |
0,73 |
0,83 |
0,78 |
Московская бойцовая |
11 |
20,81 |
5,62 |
1,00 |
0,75 |
0,84 |
0,80 |
П р и м е ч а н и е. Н1 — средняя гетерозиготность (11), Н2 — скорректированная средняя гетерозиготность (11), Н3 — средняя гетерозиготность по Jin & Chakraborty (12). |
|||||||
Выбор исследуемых пород, как упоминалось выше, был обусловлен наличием у них крови бойцовой птицы. Сравнительное исследование узбекской бойцовой породы и 29 популяций кур разного происхождения, выполненное по морфологическим признакам, показало, что орловская порода находится с ней в одном кластере кладограммы (9). Происхождение орловской ситцевой породы кур не определено, хотя у нее отмечаются некоторые морфологические признаки бойцового типа (см. табл. 1). Подобный анализ по промерам и частоте аллелей биохимических локусов выявил относительную близость кур узбекской бойцовой и московской бойцовой пород (10). Наблюдаемое сходство юрловских голосистых и московских бойцовых кур убедительно объяснить пока трудно, но полученные факты позволяют предположить генетическое пересечение этих пород в прошлом.
В перспективе при развитии методов геномной оценки, позволяющей вычленить молекулярные маркеры, связанные с определенными признаками, появится возможность переносить в процессе селекции полиморфные гены, контролирующие желательные качества (крепость ног, тип поведения, резистентость к заболеваниям и др.) (13).
Таким образом, в процессе длительного формирования изученные породы дивергировали, что в наибольшей степени проявилось у кур орловской породы. В то же время между ними обнаружено генетическое сходство, которое можно объяснить принадлежностью в прошлом к корню бойцовых кур.
Л И Т Е Р А Т У Р А
1. С е м е н о в а С.К., Ф и л е н к о А.Л., В а с и л ь е в В.А. Использование полиморфных маркеров ДНК для идентификации пород кур различного происхождения. Генетика, 1996, 32(5): 795-803.
2. T a d a n o R., N i s h i b o r i M., N a g a s a k a N., T s u d z u k i M. Assessing genetic diversity and population structure for commercial chicken lines based on forty microsatellite analyses. Poult. Sci., 2007, 86(11): 2301-2308.
3. Т ы щ е н к о В.И., Д е м е н т ь е в а Н.В., Т е р л е ц к и й В.П., Я к о в л е в А.Ф. Оценка генетического разнообразия в популяциях кур на основе геномной дактилоскопии. С.-х. биол., 2002, 6: 43-46.
4. Т ы щ е н к о В.И., М и т р о ф а н о в а О.В., Д е м е н т ь е в а Н.В., Т е р л е ц
к и й В.П., Я к о в л е в А.Ф. Оценка генетического разнообразия в породах и экспериментальных популяциях кур с помощью ДНК-фингерпринтинга. С.-х. биол., 2007, 4: 29-33.
5. L o y w i c k V., B e d ' h o m B., P i n a r d - v a n d e r L a a n M., P i t e l F., V e r r i e r E., B i j m a P. Evolution of the polymorphism at molecular markers in QTL and non-QTL regions in selected chicken lines (оpen access publication). Genet. Sel. Evol., 2008, 40: 639-661.
6. T a d a n o R., N i s h i b o r i M., I m a m u r a Y., M a t s u z a k i M., K i n o s h i t a K., M i z u t a n i M., N a m i k a w a T., T s u d z u k i M. High genetic divergence in miniature breeds of Japanese native chickens compared to Red Jungle Fowl, as revealed by microsatellite analysis. Anim. Genet., 2008, 39(1): 71-78.
7. Я к о в л е в А., Д е м е н т ь е в а Н., Т е р л е ц к и й В., Т ы щ е н к о В., М и т
р о ф а н о в а О.В., Т у ч е м с к и й Л., Г л а д к о в а Г. Контроль генетического разнообразия при использовании ДНК-технологии. Птицеводство, 2008, 12: 3-5.
8. M a t h u r P.K., P o n s u k s i l i S., G r o e n A.F. Estimation of genetic variability within and between populations using DNA fingerprinting. Proc. of the 5th World Congress on genetics applied to livestock production. Guelph, Canada, 1994: 528-531.
9. М о и с е е в а И.Г., С е м е н о в а С.К., Г о р б а ч е в а Н.С. и др. Генетическая структура и происхождение старой русской орловской породы кур. Генетика, 1994, 30(5): 681-694.
10. M o i s e y e v a I.G., R o m a n o v M.N., N i k i f o r o v A.A., S e v a s t y a n o v a A.A., S e m y e n o v a S.K. Evolutionary relationship of Red Jangle Fowl and chicken breeds. Genet. Select. Evol., 2003, 35: 403-423.
11. S t e p h e n s J.C., G i l b e r t D.A., Y u h k i N., O ’ B r i e n S.J. Estimation of heterozygosity for single-probe multilocus DNA fingerprints. Mol. Biol. Evol., 1992, 9: 729-743.
12. R o g s t a d S.H., P e l i c a n S. GELSTATS: a computer program for population genetics analysis using VNTR multilocus probe data. BioTechniques, 1996, 21: 1128-1131.
13. C h a n E.K.K., H a w k e n R., R e v e r t e r A. The combined effect of SNP-marker and phenotype attributes in genome-wide association studies. Anim. Genet., 2008, 40(2): 149-156.
A.F. Yakovlev, V.I. Tyshchenko, N.V. Dement’eva, O.V. Mitrofanova, V.P. Terletskii, A.B. Vakhrameev
By the method of DNA fingerprinting the authors determined the intra- and interbreed coefficients of genetic relation, genetic distances and the degree of heterogeneity in hens of the Yurlovskaya loud-voiced, Orlovskaya calico, Uzbekskaya fighting and Moskovskaya fighting breeds, having the genetic roots of fighting hens. The Orlovskaya calico breed is the most genetic distant from three other studied breeds. The level of heterozygosity in the hens of this breed is also the lowest. In studied breed the authors revealed the marker fragments of DNA (or their absence). The problems of genetic interrelation between four studied breeds are discussed in connection with their belonging to fighting type.
Keywords: DNA polymorphism, DNA fingerprinting, breeds of chickens, genetic distance, heterozygosity.
ГНУ Всероссийский НИИ генетики и разведения |
Поступила в редакцию |
Оформление электронного оттиска
![]()
![]()




