doi: 10.15389/agrobiology.2020.3.597rus
УДК 633.2.03:574.4:581.192(470.67)
Работа выполнена при поддержке программы Президиума РАН №1.21П «Биоразнообразие природных систем. Биологические ресурсы России: оценка состояния и фундаментальные основы мониторинга».
О БАЛАНСЕ КАЛЬЦИЯ В ТРАВЯНЫХ ЭКОСИСТЕМАХ ТЕРСКО-КУМСКОЙ НИЗМЕННОСТИ ПРИКАСПИЯ
Г.Н. ГАСАНОВ1, 2, Т.А. АСВАРОВА1, К.М. ГАДЖИЕВ1, Р.Р. БАШИРОВ1, З.Н. АХМЕДОВА1, А.С. АБДУЛАЕВА1,
Ш.К. САЛИХОВ1, Н.И. РАМАЗАНОВА1, А.Ш. ГИМБАТОВ2, М.Р. МУСАЕВ2, Н.Р. МАГОМЕДОВ2,3, Р.З. УСМАНОВ1
Терско-Кумская низменность занимает Северо-Западную часть Прикаспийской низменности. Почвенно-растительный покров территории определяется аридностью климата с частыми ветрами, наличием легких по гранулометрическому составу и засоленных почв, высокой пастбищной нагрузкой. В настоящем исследовании впервые дифференцированно определены продуктивность пастбищ Терско-Кумской низменности по структуре фитомассы (фотосинтезирующая масса, ветошь, степной войлок, корни), содержание и запасы кальция в благоприятный (апрель) и засушливый (август) периоды по сезонам года, рассчитан баланс Са в фитоценозах основных типов почв. Целью работы было определение объемов накопления, распределения и запасов кальция в структуре фитомассы для основных типов почв Терско-Кумской низменности. Исследования проводили на Кочубейской биосферной станции Прикаспийского института биологических ресурсов Дагестанского научного центра РАН в 2011-2016 годах. Объектами исследований были травяные фитоценозы на светло-каштановой и лугово-каштановой почвах и солончаке типичном автоморфном. Учет накопления растительного вещества за единицу времени на единице площади, содержания Са по элементам структуры (зеленая масса, ветошь, степной войлок, корни), а также расчет баланса Са в экосистемах выполняли по А.А. Титляновой с соавт. (1988). Содержание Са в растениях определяли при помощи системы капиллярного электрофореза Капель-105М («Люмэкс», Россия) со специализированным программным обеспечением Эльфоран («Люмэкс», Россия). Показано, что наибольшее количество фитомассы накапливалось на светло-каштановой почве. На лугово-каштановой почве фотосинтезирующей массы, ветоши, степного войлока и корней накапливалось меньше соответственно в 2,3, 1,5, 2,3 и 2,2 раза; на солончаке типичном — в 2,6, 1,7, 2,5 и 2,7 раза, чем на светло-каштановой. Возможно, это связано со снижением проективного покрытия с 77,0 % на светло-каштановой почве до 48,5 % на лугово-каштановой и 43,5 % на солончаке. В видовом составе фитоценозов на светло-каштановой почве преобладали представители Poaceae (51 %), на лугово-каштановой — Asteraceae (30 %), на солончаке типичном — также Asteraceae (17 %). Масса корней на лугово-каштановой почве и солончаке типичном была в 2,2 и 2,9 раза меньше, чем на светло-каштановой почве. Их доля в суммарном урожае фитомассы по типам почв колебалась от 85,0 до 87,2 %. В зеленой фитомассе содержание Са по сезонам года и типам почв находилось в пределах 0,40-0,48 %, в ветоши — 0,50-054, в войлоке — 1,00-1,31, в подземных органах — 1,14-1,38 %. По типам почв оно снижалось в ряду светло-каштановая > лугово-каштановая > со-лончак в связи с динамикой рН (8,6 > 8,2 > 8,0), увеличением степени засоления и смещением химизма засоления от сульфатного типа в сторону сульфатно-хлоридного. Запасы Са в надземной массе фитоценоза за вегетационный период на светло-каштановой почве составляли 2,32 кг/га·в год и превышали аналогичные показатели на лугово-каштановой почве и солончаке типичном соответственно в 2,7 и 3,1 раза. Его запасы в корневой массе по всем типам почв были больше, чем в надземной, в 12,6 раза. Из потребленного из почвы Са при разложении степного войлока обратно в светло-каштановую почву возвращалось 42,0 %, подземных органов — 58,0 %; в лугово-каштановую почву — соответственно 36,0 и 64,0 %; в солончак типичный — 1,1- и 2,3-кратное количество. Выявлено, что различие в динамике накопления Са в структуре фитомассы полупустынных фитоценозов (фотосинтезирующая масса, ветошь, степной войлок, корни) зависит от видового состава, типа почвы и сезона года.
Ключевые слова: фитоценоз, накопление фитомассы, транслокация растительного вещества, концентрация Ca, запасы Ca, бюджет кальция.
Для Терско-Кумская низменности (Северо-Западную часть Прикаспийской низменности) характерны легкие по гранулометрическому составу и засоленные почвы, аридность климата, высокая пастбищная нагрузка, деградированность почвенного и растительного покрова. Почвы с низким содержанием гумуса и основных элементами питания растений в сочетании с неблагоприятным водно-солевом режимом имеют низкую продуктивность (1). Однако эти оценки получены только для фотосинтезирующей части фитомассы, в то время как в структуре фитоценоза присутствуют и другие элементы — ветошь, степной войлок и корневая масса.
Исследования элементного состава фитоценозов Терско-Кумской низменности проводили многие авторы. В частности, имеются данные по относительному содержанию кальция в вегетирующей воздушно-сухой массе (сене) (2, 3), но нет аналогичных сведений с учетом структуры растительного покрова (зеленая масса, ветошь, войлок, корни). Отсутствие этих данных не позволяет судить о запасах биофильных элементов, в том числе Са, и оценить его баланс в травяных экосистемах, в том числе дифференцированно по основным типам почв.
На территории Терско-Кумской низменности содержание Са растениях больше, чем на прилегающей Присулакской низменности, — 2,014±0,02 против 1,623±0,25 г/кг (4). При этом отмечают постепенное снижение его накопления с ноября по апрель следующего года. Г.И. Гиреев с соавт. объясняют это трансформацией лугово-полынных сообществ в эфемеро-полынные и полынные. Можно предположить, что такое снижение количества Са в фитоценозах имеет другую причину и связано с транслокацией веществ между элементами структуры фитоценоза при смене фаз развития растений.
Отмечают, что решающее влияние на накопление химических элементов в растениях оказывает увлажненность, в частности содержание Са повышается по мере увеличения количества осадков (5, 6). Также выявлено, что благоприятные гидротермические условия в период вегетации растений служат фактором, способствующим повышению содержания Са в почве (7).
В настоящем исследовании в условиях Терско-Кумской полупустыни для основных типов почв дифференцированно (с учетом структуры фитомассы) оценена продуктивность травяных экосистем в связи с запасами Са в наиболее благоприятный по климатическим условиям (апрель) и засушливый (август) периоды года, а также рассчитан баланс Са в экосистемах.
Цель работы — изучить аккумуляцию кальция в растительном веществе с учетом его структуры и распределение этого элемента в травяных экосистемах на основных типах почв Терско-Кумской низменности.
Методика. Исследования проводили на Кочубейской биосферной станции Прикаспийского института биологических ресурсов Дагестанского научного центра РАН (ПИБР ДНЦ РАН, КБС, 44.40720 с.ш., 46.24771 в.д) в 2011-2016 годах. Содержание, запасы и бюджет кальция с учетом сезонов года (апрель, август) детально анализировали в 2013-2015 годах. Изучали травяные фитоценозы на светло-каштановой и лугово-каштановой почвах и солончаке типичном автоморфном Терско-Кумской низменности. Содержание гумуса в слое 0-30 см светло-каштановой почвы составляло 1,12 %, Р2О5 — 1,11 мг/100 г, К2О — 20,12 мг/100 г почвы, плотность — 1,14 г/см3, наименьшая влагоемкость — 20,6 %; для лугово-каштановой эти показатели составляли соответственно 0,54 %, 0,7 мг/100 г, 18,2 мг/100 г, 1,42 г/см3, 26,7 %; для солончака типичного автоморфного — 0,48 %; 0,66 мг/100 г; 16,8 мг/100 г; 1,45 г/см3 и 30,3 %. Тип засоления менялся от хлоридно-сульфатного до хлоридного в зависимости от гидротермических условий сезонов года, степень засоления — от слабой до очень сильной.
Учет накопления растительного вещества за единицу времени на единице площади, содержания Са в зеленой массе, ветоши, степном войлоке и корнях, а также расчет баланса Са в экосистемах проводили по А.А. Титляновой (7). Содержание Са в растительном веществе определяли методом капиллярного электрофореза (система Капель-105М, «Люмэкс», Россия) в режиме определения катионов и анионов со специализированным программным обеспечением Эльфоран («Люмэкс», Россия) (8). Всего проанализировали 150 проб в 3 биологических и аналитических повторностях.
Латинские названия видов приведены по Р.А. Муртазалиеву (9).
Статистическую обработку данных проводили в программе Microsoft Exсel 2010. Рассчитывали средние величины (M) и ошибки среднего (±SEM), уравнения линейной регрессии, коэффициент корреляции (r), коэффициент детерминации (R).
Результаты. В условиях равнины наиболее продуктивны фитоценозы на светло-каштановой почве: в среднем за 2011-2016 годы накопление воздушно-сухой фитомассы здесь было в 2,2 раза выше, чем в случае лугово-каштановой почвы. На солончаке типичном автоморфном продуктивность фитоценоза снижалась в 2,6 раза по сравнению с таковой для светло-каштановой почвы (табл.).
На лугово-каштановой почве и солончаке типичном накапливалось столько же ветоши (соответственно 102,8 и 100,5 %), сколько и зеленой массы, а на светло-каштановой почве — на 33,7 % меньше, что было связано с видовым составом фитоценозов.
На светло-каштановой почве доля эфемеров из семейства мятликовых, которые были представлены полевичкой малой (Eragrostic minor Host.), мятликом луковичным (Poa bulbosa L.), мортуком восточным (Eremopyrum orientale L.), составляла 51,4 % по количеству и 19,6 % по массе. На лугово-каштановой почве и солончаке фитомасса полыни таврической (Artemisia taurica Willd)и полыни Лерха (A. lercheana Web. ex Stechm) оставалась достаточно высокой (37,7 % по количеству и 83,7 % по массе), поскольку их ветошь за апрель-август не успевала полностью перейти в степной войлок.
По данным многих авторов, вклад подземных органов в общую массу травянистых фитоценозов составляет 50-90 % (10, 11). В наших исследованиях масса корней на лугово-каштановой почве и солончаке типичном оказалась в 2,2 и 2,9 раза меньше, чем на светло-каштановой почве. Их доля в суммарном урожае фитомассы по типам почв колебалась от 85,0 до 87,2 %. Такое высокое соотношение надземной и подземной массы (1:6,8-1:5,7) характерно для всех засушливых регионов мира (12, 13).
По нашим данными, в условиях полупустыни связь между накоплением надземной (х) и подземной (у) массы у растений в экосистемах в зависимости от типа почв описывали следующие уравнения регрессии:
на светло-каштановой y = 0,6935x + 73,7823 (r = 0,97, R = 0,96);
на лугово-каштановой y = 1,1804x + 30,7254 (r = 0,96, R = 0,95);
на солончаке типичном y = 1,7925x + 21,0415 (r= 0,97, R = 0,95).
Основной причиной снижения сборов фитомассы на лугово-каш-тановой почве и солончаке типичном становится усиление засоления почвы и изменение его характера в сторону увеличения соотношения ионов Cl-:SO42- (11). Следует отметить, что продуктивность травяных экосистем Терско-Кумской низменности была значительно ниже, чем в других регионах страны, где она составляет 15-20 т/га (10, 11). Только на светло-каштановой почве этот показатель приближался к нижнему из указанных пределов, а на лугово-каштановой почве и солончаке типичном оказался в 2,2-2,6 раза ниже. Аналогичные результаты получены другими авторами в районе проведения наших исследований (5), в Астраханской области (14), в условиях сухой и опустыненной степи Тувы (12, 13), в засушливых районах Ирана (15), Панамы (5), Китая (16), в регионах с аридным климатом.
В жизни растений Са играет важную роль в образовании клеточных оболочек, принимает участие в поддержании структуры хромосом, митохондрий и рибосом (17, 18), повышает солевую толерантность (19-21), устойчивость растений к тепловому стрессу (22) и негативному воздействию тяжелых металлов (23), оптимизирует рост и развитие корней (24).
Средние за 2013-2015 годы (апрель, август) показатели содержания Са в зеленой массе и других элементах структуры растительного покрова в целом соответствовали накопленному урожаю фитомассы и составляли на светло-каштановой почве 0,48 %, на лугово-каштановой — 0,41 %, на солончаке — 0,40 %. Были отмечены существенные различия в содержании элемента по видам растений. На светло-каштановой почве 51 % от суммарного проективного покрытия (77 %) приходился на злаковые — костер кровельный Anisantha tectorum (L.) Nevski, костер растопыренный Bromus squarrosus L., мятлик луковичный Poa bulbosa L., полевичку малую Eragrostis minor Host и мортук пшеничный Eremopyrum triticeum (Gaertn.) Nevski, 15 % — на полыни Artemisiataurica Willd и A. lercheana Web. ex Stechm, 5 % — на солянки, включая рогач песчаный Ceratocarpus arenarius L., солянку грузинскую Salsolaiberica Sennen & Pau, петросимонию раскидистую Petrosimoniabrachiata (Pall.) Bunge, 3 % — на гвоздичные (смолевка коническая Silene conicaL. и грыжник седой Herniaria incana Lam.) и 3 % — на крестоцветные (бурачок пустынный Alussum desertorum Stapf.). Содержание кальция у сложноцветных (виды полыни) составляло 1,03 %, у маревых (солянки) — 0,93 %, у гвоздичных — 0,38 %, у крестоцветных — 0,22 % и у злаковых — 0,58 %
На лугово-каштановой почве проективное покрытие мятликовыми снижалось в 5,1 раза, гвоздичными — в 2,0 раза, крестоцветными — в 1,5 раза, тогда как полынями из семейства сложноцветных — увеличивалось в 2,0 раза. Несмотря на то, что на этом типе почв доля сложноцветных и маревых с высоким накоплением Са повышалась, среднее содержание элемента в группе мятликовых снижалось по сравнению с показателями, полученными на светло-каштановой почве, до 0,47 %, у маревых — до 0,90 % при незначительных отклонениях в фитомассе у других семейств. На солончаке типичном общее проективное покрытие фитоценозом (43,5 %) приближалось к показателю на лугово-каштановой почве (48,5 %), но содержание элемента снижалось только у солянок (на 0,24 %) при относительно близких значениях в других группах растений.
Уменьшение содержания Са в фитомассе на почвах в ряду светло-каштановая > лугово-каштановая > солончак мы связываем с незначительным понижением рН почвенного раствора в этом же ряду (8,6 > 8,2 > 8,0), что становится причиной более легкой доступности Са из почв, то же подтверждают и другие авторы (25). Кроме того, сказывается усиление засоления и переход от его сульфатного типа в сторону сульфатно-хлоридного (11). Эти факторы существенно снижают доступность Са для растений из лугово-каштановой почвы и солончака типичного по сравнению со светло-каштановым типом.
Известно, что поступление ионов различных металлов из почвы в корневую систему и надземные органы растений осуществляется за счет динамики осмотического давления с участием переносчиков, кодируемых различными генами (26). Роль типа почвы (х) и видового состава фитоценозов (y) в изменении концентрации Са (z) описывало следующее уравнение: z = 0,1057 - 0,4999x + 1,1898y.
В ветоши и войлоке содержание Са выравнивалось, а в подземных органах снижалось от 1,38 % на двух первых типах почв до 1,14 % на солончаке типичном (рис. 1). Достаточно высокое количество Са в фитомассе в условиях непромывного водного режима способствовало поддержанию в почвенном растворе нейтральной и слабощелочной реакции (10).
Запасы элемента в корневой массе на всех почвах были больше, чем в надземной, в среднем в 12,6 раза (73,81 кг/га против 5,85 кг/га). С учетом приведенных данных был составлен баланс Са в биогеоценозах рассматриваемых типов почв (рис. 2). В травяных экосистемах кальция потребляется значительно больше, чем в лесных (16). В наших исследованиях на светло-каштановой почве из потребленных фитоценозом 125,23 кг/га Са в год при разложении степного войлока в почву возвращалось 42,0 %, подземных органов — 58,0 %. На лугово-каштановой почве доля Са, поступившего при разложении войлока, снижалась до 36,8 %, на солончаке типичном — увеличивалась до 48,4 %.
Относительно высокая продуктивность фитоценозов на светло-каштановой почве способствует обогащению ее верхних горизонтов Са и другими биофильными элементами (Fe, Si, Al), которые служат основным фактором биосинтеза вторичных глинистых минералов, формирующих поглощающий комплекс почвы. Высокое содержание в фитомассе травянистых растений Са (наряду Mg и S) приводит к насыщению поглощающего комплекса почв Терско-Кумской низменности этим сильным коагулятором, обеспечивающим формирование благоприятных агрофизических свойств почвы и улучшение ее структуры (1).
На всех типах почв весной наблюдалось более высокое содержание Са по сравнению с летним периодом (I декада августа), что объясняется благоприятным водным режимом почвы в этот период, способствующим поступлению химических элементов в растения. Снижение количества Са в августе связано с увеличением засоленности почвы, ростом осмотического давления почвенного раствора и замедлением поступления почвенной влаги и питательных элементов в растения. Эти данные подтверждают справедливость вывода о том, интенсивность поступления химических элементов, включая Са, в растения связана с благоприятными гидротермическими условиями (10), а не с трансформацией лугово-полынных сообществ в эфемеро-полынные и полынные (3, 4).
Еще одна причина снижения накопления Са в пастбищных фитоценозах к концу вегетационного периода или к весне следующего года — отток части элементов из стареющих и отмирающих растительных тканей в молодые или вновь формируемые. По мнению А.А. Титляновой (10), процесс ретранслокация питательных элементов универсален и присущ всем экосистемам. Следовательно, на основных типах изученных почв достаточное содержание Са в фитомассе, а в последующем его поступление в верхнюю часть почвенного профиля становится основой биологических циклов Са в травяных ценозах.
Таким образом, максимальные запасы фотосинтезирующей фитомассы (4,84 кг·га-1·год-1), ветоши (3,21 кг·га-1·год-1), степного войлока (4,32 кг·га-1·год-1) и корней (83,71 кг·га-1·год-1) на Терско-Кумской низменности накапливались на светло-каштановой почве. На лугово-каштановой почве эти показатели снижались соответственно в 2,3; 1,5; 2,3 и 2,2 раза, на солончаке типичном — в 2,6; 1,7; 2,5 и 2,7 раза. Накопление Са в надземной и подземной массе соответствовала урожайности фитоценозов. В зеленой массе его содержание составляло в среднем 0,48-0,40 %, в войлоке — 1,0-1,3 %, в подземных органах — 1,14-1,38 %. Максимальные запасы элемента в надземной массе растений за вегетационные периоды накапливались на светло-каштановой почве (9,71 кг·га-1·сут-1), превышая аналогичные показатели на лугово-каштановой почве и солончаке типичном соответственно в 2,7 и 3,1 раза. На светло-каштановой почве при разложении степного войлока в почву возвращалось 42,0 % потребленного Са, на лугово-каштановой — 36,0 %, на солончаке типичном – 48,4 %. Такое же его количество перемещалось в надземные органы. При разложении подземных органов в растения поступало соответственно по типам почв 58,0; 64,0 и 51,6 % Са. Выявлено, что различие в динамике накопления Са в структуре фитомассы (фотосинтезирующая масса, ветошь, степной войлок, корни) полупустынных фитоценозов зависит от видового состава, типа почвы, сезона года.
Накопление воздушно-сухой надземной и подземной фитомассы в структурных элементах растительного покрова и аккумуляция Са для разных типов почвы Терско-Кумской низменности (Кочубейская биосферная станция ПИБР ДНЦ РАН, 2011-2016 годы) |
||||
Растительное вещество |
Тип почвы |
Фитомасса, т/га в год (M±SEM) |
Са |
|
содержание, % (M±SEM) |
запасы, кг/га |
|||
Зеленая масса |
Светло-каштановая |
4,84±0,02 |
0,48±0,02 |
2,32 |
Лугово-каштановая |
2,11±0,01 |
0,41±0,01 |
0,86 |
|
Солончак типичный |
1,89±0,03 |
0,40±0,03 |
0,76 |
|
Ветошь |
Светло-каштановая |
3,21±0,02 |
0,54±0,03 |
1,73 |
Лугово-каштановая |
2,17±0,01 |
0,50±0,01 |
1,08 |
|
Солончак типичный |
1,91±0,04 |
0,50±0,02 |
0,96 |
|
Войлок |
Светло-каштановая |
4,32±0,03 |
1,31±0,01 |
5,66 |
Лугово-каштановая |
1,89±0,02 |
1,30±0,02 |
2,46 |
|
Солончак типичный |
1,71±0,02 |
1,00±0,01 |
1,71 |
|
Подземная масса |
Светло-каштановая |
83,71±0,03 |
1,38±0,02 |
115,52 |
Лугово-каштановая |
38,19±0,03 |
1,38±0,03 |
52,70 |
|
Солончак типичный |
31,19±0,02 |
1,14±0,03 |
35,56 |
|
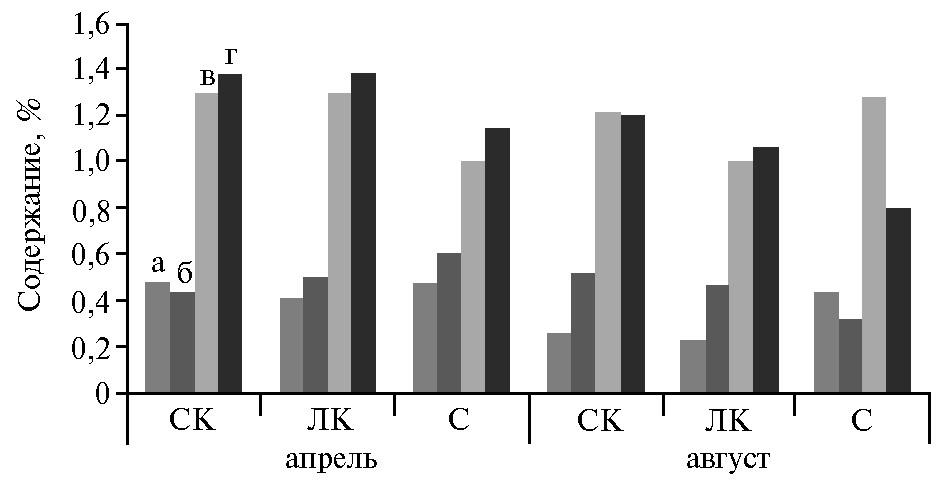
Рис. 1. Накопление Са в фитоценозах Терско-Кумской низменности в зависимости от типа почв и сроков определения: СК — светло-каштановая почва, ЛК — лугово-каштановая почва, С — солончак типичный; а — зеленая масса, б — ветошь, в — войлок, г — подземные органы (Кочубейская биосферная станция ПИБР ДНЦ РАН, 2013-2015 годы).
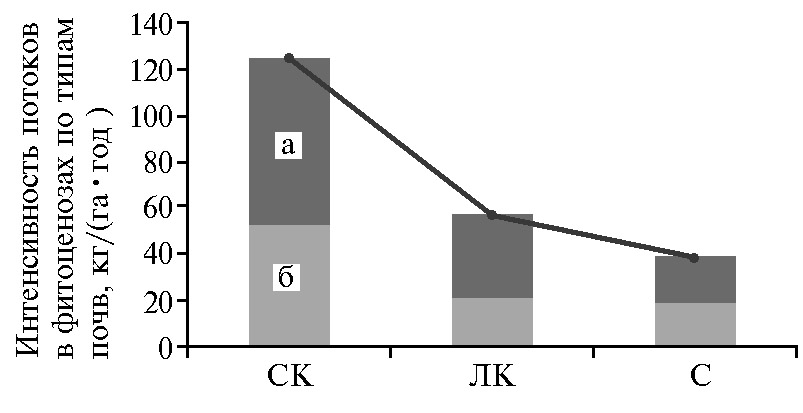
Рис. 2. Бюджет кальция в фитоценозах Терско-Кумской низменности в зависимости от типа почв: СК — светло-каштановая почва, ЛК — лугово-каштановая почва, С — солончак типичный; а — поступило в почву при разложении подземных органов, б — поступило в почву при разложении войлока; кривая — потреблено из почвы (Кочубейская биосферная станция ПИБР ДНЦ РАН, 2013-2015 годы).
ЛИТЕРАТУРА
- Баламирзоев М.А., Мирзоев Э.М.-Р., Аджиев А.М., Муфараджев К.Г. Почвы Дагестана. Экологические аспекты их рационального использования. Махачкала, 2008.
- Усманов Р.З., Осипова С.В., Джалалова М.И., Бабаева М.А. Использование методов фитомелиорации на деградированных пастбищах Терско-Кумской низменности. Юг России: экология, развитие, 2008, 3(3): 109-111.
- Загидова Р.М., Бийболатова З.Д., Асгерова Д.Б., Абдурашидова П.А. Экология смен в растительных группировках Терско-Кумской и Терско-Сулакской низменностей. Вестник Дагестанского государственного университета, 2015, 30(1): 60-65.
- Гиреев Г.И., Луганова С.Г., Салихов Ш.К. Приоритетные макроэлементы в растительности пастбищ регионов Дагестана. Вестник Тамбовского университета. Серия: Естественные и технические науки, 2013, 18(3): 767-770.
- Santiago LS., Schuur EA., Silvera K. Nutrient cycling and plant-soil feedbacks along a precipitation gradient in lowland Panama. Journal of Tropical Ecology, 2005, 21(4): 461-470 (doi: 10.1017/s0266467405002464 ).
- Dijkstraa F.A., Heb M., Johansenc M.P., Harrisonc J.J., Keitel C. Plant and microbial uptake of nitrogen and phosphorus affected by drought using 15N and 32P tracers. Soil Biology and Biochemistry, 2015, 82: 135-142 (doi: 10.1016/j.soilbio.2014.12.021).
- Титлянова А.А., Базилевич Н.И., Снытко В.А., Дубынина С.С., Копотева Т.А., Магомедова Л.Н., Миронычева-Токарева Н.П., Нефедьева Л.Г., Семенюк Н.В., Тишков А.А., Ти Тран, Хакимзянова Ф.И., Шатохина Н.Г., Шмакова Е.И. Биологическаяпродуктивностьтравяныхэкосистем. Географические закономерности и экологические особенности /Под ред. В.Б. Ильиной. Новосибирск, 1988: 109-127.
- Комарова Н.В., Каменцев Я.С. Практическое руководство по использованию систем капиллярного электрофореза «КАПЕЛЬ». СПб, 2006.
- Муртазалиев Р.А. Конспект флоры Дагестана /Под ред. Р.В. Камелина. Махачкала, 2009.
- Титлянова А.А. Бюджет элементов питания в экосистемах. Почвоведение, 2007, 12: 1422-1429.
- Гасанов Г.Н., Асварова Т.А., Гаджиев К.М., Ахмедова З.Н., Абдулаева А.С., Баширов Р.Р., Арсланов М.А. Состояние и приемы восстановления продуктивности растительного покрова Терско-Кумской полупустыни. Известия Самарского научного центра РАН, 2016, 18(2): 59-64.
- Курбатская С.С. Изменение структуры и запаса растительного вещества на мониторинговых участках степных экосистем. Новые исследования Тувы, 2009, 4: 209-224.
- Титлянова А.А., Самбуу А.Д. Детерминированность и синхронность залежной сукцессии в степях Тувы. Известия РАН. Серия биологическая, 2014, 6: 621-630.
- Тютюма Н.В., Булахтина Г.К. Проблема опустынивания аридной зоны Астраханской области в условиях изменения климата и повышенного антропогенного воздействия. Труды института геологии Дагестанского научного центра РАН, 2016, 1: 68-72.
- Saffariha M., Azarnivand H., Tavili A. Effects of grazed exclosure on some of nutrient elements of aerial and underground organs of Artemisia sieberi, Stipa hohenacheriana and Salsola rigida. International Journal of Agronomy and Agricultural Research, 2014, 4(2): 62-70.
- Zuo X.A., Knops J.M.H., Zhao X.Y., Zhao H.L., Li Y.Q., Guo Y.R. A positive correlation between plant diversity and productivity is indirectly caused by environmental factors driving spatial pattern of vegetation composition in semiarid sandy grassland. Biogeosciences Discussions, 2011, 8: 1795-1801.
- Marschner's mineral nutrition of higher plants. 3rd edition /P. Marschner (ed.). Amsterdam, Netherlands, Elsevier/Academic Press, 2011 (doi: 10.1017/s001447971100130x).
- Shen J., Yuan L., Zhang J., Bai Z., Chen X., Zhang W., Zhang F. Phosphorus dynamics: from soil to plant. Plant Physiology, 2011, 156(3): 997-1005 (doi: 10.1104/pp.111.175232).
- Khan M.N., Siddiqui M.H., Mohammad F., Naeem M. Interactive role of nitric oxide and calcium chloride in enhancing tolerance to salt stress. Nitric Oxide, 2012, 27(4): 210-218 (doi: 10.1016/j.niox.2012.07.005).
- Hadi M.R., Karimi N. The role of calcium in plants' salt tolerance. Journal оf Plant Nutrition, 2012, 35(13): 2037-2054 (doi: 10.1080/01904167.2012.717158).
- Manishankar P., Wang N., Köster P., Alatar A., Kudla J. Calcium signaling during salt stress and in the regulation of ion homeostasis. Journal of Experimental Botany, 69(17): 4215-4226 (doi: 10.1093/jxb/ery201).
- Tan W., Meng Q.W., Brestic M., Olsovska K., Yang X. Photosynthesis is improved by exogenous calcium in heat-stressed tobacco plants. Journal of Plant Physiology, 2011, 168(17): 2063-2071 (doi: 10.1016/j.jplph.2011.06.009).
- Huang D., Gong X., Liu Y., Zeng G., Lai C., Bashir H., Zhou L., Wang D., Xu P., Cheng M., Wan J. Effects of calcium at toxic concentrations of cadmium in plants. Planta, 2017, 245(5): 863-873 (doi: 10.1007/s00425-017-2664-1 ).
- Zhang T., Yang J., Sun Y., Kang Y., Yang J., Qi Z. Calcium deprivation enhances non-selective fluid-phase endocytosis and modifies membrane lipid profiles in Arabidopsis roots. Journal of Plant Physiology, 2018, 226: 22-30 (doi: 10.1016/j.jplph.2018.04.002).
- González-Fontes A., Navarro-Gochicoa M.T., Ceacero C.J., Herrera-Rodríguez M.B., Camacho-Cristóbal J.J., Rexach J. Chapter 9 — Understanding calcium transport and signaling, and its use efficiency in vascular plants. In: Plant macronutrient use efficiency. Molecular and genomic perspectives in crop plants /M.A. Hossain, T. Kamiya, D.J. Burritt, L.-S.P. Tran, T. Fujiwara (eds.). Academic Press, 2017: 165-180 (doi: 10.1016/B978-0-12-811308-0.00009-0).
- Krämer U., Talke I.N., Hanikenne M. Transition metal transport. FEBS Letters, 2007, 581(12): 2263-2272 (doi: 10.1016/j.febslet.2007.04.010).