doi: 10.15389/agrobiology.2019.3.512rus
УДК 633.18:581.1:631.547
РЕГУЛЯЦИЯ ОТТОКА АССИМИЛЯТОВ В ЗЕРНОВКИ ДЛЯ ПОВЫШЕНИЯ ПРОДУКТИВНОСТИ ПОСЕВОВ РИСА (Oryza sativa L.) ПРИ ВЫРАЩИВАНИИ У СЕВЕРНОЙ ГРАНИЦЫ АРЕАЛА КУЛЬТУРЫ
А.Х. ШЕУДЖЕН1, 2, Т.Н. БОНДАРЕВА1, 2, П.Н. ХАРЧЕНКО3, И.А. ДОРОШЕВ2
Рис в основном выращивают в странах с благоприятным для культуры климатом, лежащих между экватором и 45° широты, но в последние годы интерес к расширению географии рисосеяния в северном направлении возрастает. В России зона рисоводства Краснодарского края, где культура возделывается давно и достаточно успешно, находится у северной границы ее ареала. По этой причине в некоторые годы созревание риса совпадает с неблагоприятными погодными условиями, удлиняющими вегетационный период, что сдвигает время уборки на более поздние сроки и может приводить к потерям значительной части урожая. В связи с этим необходимы приемы, позволяющие физиологически ускорить созревание растений без снижения их продуктивности. Один из них — сеникация (лат. senium — старение), то есть целенаправленное регулирование метаболизма на завершающих стадиях развития растений. Сеникацию следует применять при задержке созревания, что чаще всего наблюдается на поздних (вследствие переноса сроков сева из-за погодных условий), изреженных или получивших избыточное азотное питание посевах. Хотя в настоящее время сеникация посевов риса используется мало, что обусловлено главным образом продолжительным теплым периодом в многолетнем цикле колебаний температур, вероятность наступления в ближайшие годы периода с неблагоприятными для созревания и уборки риса погодными условиями высока. Эффективность метода зависит от состава сениканта, срока проведения и погодных условий. Кроме того, для применения сеникации на современных сортах риса необходимо определить эффективность и регламент проведения сеникации с учетом особенностей процесса созревания зерновок. В представленной работе мы сравнили некоторые физиологические показатели растений при созревании зерна, элементы структуры урожая и урожайность у современного российского интенсивного сорта Хазар в зависимости от состава сеникантов и сроков их применения с учетом погодных условий за три года наблюдений. Показано, что наиболее эффективны сениканты на основе водного раствора аммонийной селитры (15 кг/га) с добавлением Mn (400 г/га) или Se (200 г/га). Модифицированный селеном раствор следует использовать при необходимости приступить к уборке через 14-20 cут. Обработка растений раствором c добавлением Mn стимулирует синтез и аттракцию ассимилятов в зерновку и сопровождается не столь быстрым завершением онтогенеза, но существенным ростом урожая. Для ускорения созревания растений риса проводят сеникацию посевов в начале фазы молочно-восковой спелости зерна. В холодные годы это позволит не только быстрее (на 5-6 сут) и значительнее (на 0,69-7,39 %), чем в контроле, снизить влажность метелок (достичь технической зрелости), но и на 4,2-9,3 % повысить урожайность. В годы с благоприятными для созревания риса погодными условиями сеникация обеспечивает не столь значительное уменьшение влажности (на 0,66-2,78 %), но большее увеличение урожайности (на 7,9-14,7 %) вследствие более быстрого и полного оттока продуктов фотосинтеза в зерновки, что сопровождается увеличением массы 1000 зерен, снижением числа стерильных и несущих щуплые зерновки колосков (пустозерность) и, как следствие, увеличением массы зерна с метелки. Исследование механизма наблюдаемых эффектов расширяет представления о биологии культуры и могут быть полезны при разработке технологий адаптации риса при смещении ареала возделывания на север с использованием сеникантов нового поколения, безопасных для человека и окружающей среды, и в программах селекции на ускоренное созревание в условиях северного рисосеяния.
Ключевые слова: Oryza sativa L., рис, сеникация, ускорение созревания, листовая подкормка, мочевина, аммонийная селитра, Mn, Se.
Рис — один из важных злаков, которым питается более половины населения Земли. В основном рис производят в странах с благоприятным для культуры климатом, лежащих между экватором и 45° широты (Южная и Юго-Восточной Азия, дающие до 90 % мирового производства риса-сырца, прежде всего Китай, Индия, Индонезия, Таиланд, Япония, а также Бразилия и США) (1, 2). В Европе рис выращивают в Испании, Италии (3, 4), Греции и Турции (5). Россия ограничивает ареал культуры с севера (42-47° с.ш.) при сосредоточении ее основных коммерческих посевов в Краснодарском крае (44-45° с.ш.) (6). В последние годы интерес к расширению географии рисосеяния возрастает. Так, по мнению исследователей, успешное выращивание риса на экспериментальных полях в юго-западной части провинции Онтарио в Канаде на территории с достаточно прохладным влажным континентальным климатом указывает на хорошие шансы этой страны стать производителем риса (7). Затопление сельскохозяйственных полей под посевы риса может также выполнять экологическую функцию, создавая водно-болотные угодья для обитания водоплавающих птиц (7) и пруды для выращивания рыбы (подобный опыт был успешным в Краснодарском крае в 1980-е годы). Проблема продвижения зоны рисосеяния на север (northerning) становится более актуальной в связи с вероятностью как глобального потепления, так и смены периода потепления похолоданием в многолетнем цикле колебания температур.
В онтогенезе растений последовательно сменяются конкурентные отношения между органами за обладание метаболитами (побег—корень, лист—цветок, лист—плод) (8, 9). На каждом его этапе существует физиологически активный центр (центр запасания), к которому направляется наи-большее количество ассимилятов. На последних стадиях развития растения конкурентные отношения в системе лист—плод складываются в пользу последнего, и пластические вещества транспортируются именно к ним. Перед формированием урожая в метаболизме листьев запускается каскад сигнальных процессов и происходит резкий сдвиг в сторону усиления гидролитических реакций, приводящих к распаду белков и крахмала на более простые соединения: аминокислоты и сахара, которые аттрагируются в генеративные органы — плоды и семена (10-12). Однако ассимиляты, как известно, аттрагируются не полностью, а удлинение периода вегетативного развития растений, зачастую имеющее место при неблагоприятных погодных условиях, сопровождается расходом ассимилятов на вегетативный рост в ущерб урожаю (13, 14).
Один из способов управления продуктивностью растения основывается на целенаправленном регулировании оттока пластических веществ из листьев и стеблей в генеративные органы. Этот агроприем, обеспечивающий ускорение созревания и повышение урожайности сельскохозяйственных культур, получил название сеникация (от лат. senium — старение). Сеникация посевов зерновых культур приводит к медленному отмиранию листьев, сходному с естественным старением (15). Механизм действия сениканта заключается в ускорении обмена веществ в тканях растений и повышении скорости оттока пластических веществ из листьев и стеблей в метелку (колос). В результате формируется более выполненное зерно, снижается пустозерность, повышается масса зерновок (16). Чем их больше было заложено в ранние периоды вегетации, тем значительнее прибавка урожая (17-20). Сеникация имеет большие перспективы на семеноводческих (особенно изреженных) посевах с выраженным кущением, так как стимулирует созревание зерновок на боковых побегах, что позволяет получить семена с меньшей матрикальной разнокачественностью и высокими посевными свойствами (21).
Разработаны теоретические и практические аспекты сеникации на посевах капусты белокочанной (22), кукурузы (16), гречихи (17), картофеля (23), подсолнечника (24), сои (25-27). Географическое положение России с ее климатическими условиями обусловливает повышенный интерес к сеникации (в отличие от других стран). В 1970-1980-е годы ее довольно часто применяли на Кубани для ускорения созревания риса. Наибольший эффект достигался при использовании водного раствора мочевины или суперфосфата с добавлением 2,4-дихлорфеноксиуксусной кислоты (2,4-Д), выполняющей роль регулятора роста. Отметим, что наряду с ускорением созревания сеникация способствовала увеличению урожайности и улучшению качества зерна риса (21).
Нами предложен физиологически обоснованный метод повышения урожайности посевов риса в условиях рискованного рисосеяния. Новизна способа состоит в разработке технологии применения новых сеницирующих смесей, так как используемые до этого технологические регламенты устарели из-за произошедшей сортосмены и запрете на присутствие в составе сеникантов аминной соли 2,4-Д (основной компонент применявшейся ранее сеницирующей смеси). Приоритет исследований закреплен за авторами патентом П. 2580162 (РФ) МКИ3 А01G 16/00. А 01 № 59/00. ФГБНУ «ВНИИ риса» (РФ): Способ сеникации посевов риса (варианты) (авторы А.Х. Шеуджен, Т.Н. Бондарева, Е.М Харитонов, И.А. Дорошев, М.А. Ладатко, Т.Х. Гиш. Заявл. 02.07.2014. Опубл. 10.04.2016. Бюл. № 10).
Цель работы заключалась в изучении динамики влажности и накопления сухого вещества растениями риса при воздействии различными сеникантами для физиологического обоснования их состава и режимов применения, обеспечивающих ускоренное созревание зерна.
Методика. Исследования проводили на полях рисовой оросительной системы ОНО ОП ЭСП «Красное» (Красноармейский р-н, Краснодарский край) в 2009-2011 годах, различающихся по гидротермическим условиям вегетационного периода. Почва участков лугово-черноземная слабовыщелоченная слабогумусная тяжелосуглинистая на аллювиальных отложениях. Использовали интенсивный среднепоздний сорт риса Хазар (создан во Всероссийском НИИ риса); предшественник — многолетние травы, посев рядовой, глубина заделки семян 1,0-1,5 см, норма высева всхожих зерен 7 млн/га; удобрения — N90P60K45, режим орошения — укороченное затопление.
Сеникацию проводили в два срока: через 10 сут после начала цветения главной метелки (молочная спелость) и в начале фазы молочно-восковой спелости зерна. Для этого растения опрыскивали водными растворами мочевины (20 кг/га) и аммонийной селитры (15 кг/га) с добавлением марганца (400 г/га) и селена (200 г/га). Марганец использовали в форме сульфата марганца, селен — селенита натрия. Норма расхода рабочего раствора 400 л/га.
Общая площадь делянки 20 м2, учетная — 10 м2, повторность 4-кратная; размещение делянок дактиль-методом.
Динамику влажности и сухой массы после сеникации определяли на каждой делянке у растений, отобранных с интервалом 7 сут в 4-кратной повторности с участков площадью по 0,25 м2 (общая площадь 1 м2).
Созревание растений оценивали по изменению влажности и сухой массы листьев, стеблей и метелок при взвешивании после 6-часового высушивания (106 °С). Урожайность зерна определяли пересчетом массы зерна с учетной площади делянки с поправкой на стандартную влажность и чистоту, массу 1000 зерен — в соответствии с ГОСТ 10842-89, пустозерность — по доле невыполненных колосков от их общего числа в метелке.
Данные представлены в виде средних (M) и стандартных ошибок средних (±SEM). Полученные результаты оценивали методом дисперсионного анализа с помощью F-критерия и критерия НСР на 5 % уровне значимости (28).
Результаты. На первом этапе мы сравнили более 50 вариантов составов сеникантов, в качестве которых испытывали макро- и микроудобрения, регуляторы роста растений, десиканты (данные не приведены). Наиболее эффективными оказались водные растворы мочевины (20 кг/га) и аммонийной селитры (15 кг/га), модифицированные марганцем (400 г/га) или селеном (200 г/га). При подборе сеникантов исходили из того факта, что, по мнению большинства исследователей, главным компонентом сениканта должен быть азот в амидной или аммонийной форме (29), и чаще всего для этих целей используют азотные удобрения: мочевину, аммонийную селитру, карбамидно-аммонийную смесь.
Влияние обработок азотом на старение листьев и урожайность культур может различаться в зависимости от сорта, дозировки, способа применения и условий выращивания (30). У риса ускорение созревания при сеникации с применением азотных удобрений связывают с действием азота, который обнаруживается в растениях уже через 30 мин после нанесения на лист. Этот азот служит дополнительным источником питания и включается в синтез органических соединений, в результате чего содержание N в листьях существенно повышается. Происходит своеобразное «переполнение» клеток листа пластическими веществами и поступающими в его ткани свободными ионами аммония, которые включаются в азотный обмен растения. В результате ускоряется биосинтез ассимилятов и их отток из вегетативных органов в зерновки. Одновременно происходит деградация хлоропластов (31, 32), ослабление фотосинтеза и интенсивности дыхания вследствие аммиачного отравления растения. По мере завершения налива зерновок неиспользованный растениями (остаточный) азот в аммонийной форме накапливается в клетках, что ингибирует функциональную активность глутаминсинтетазы и глутамин-оксоглутарат-аминотрансферазы — ферментов осуществляющих включение ионов аммония в аминокислоты и амиды (33). Причиной ускоренного завершения растениями онтогенеза считают накопление в растительных клетках аммония (29), способного вызывать в них деструктивные процессы (34, 35).
Для повышения эффективности сеникации к азотному удобрению добавляют 2,4-Д или такие важные для растения микроэлементы, как бор, селен, марганец (29, 36-38). Роль 2,4-Д заключается в облегчении проникновения иона аммония или молекулы мочевины через эпидермис в мезофилл листа, мобилизации и усилении оттока ассимилятов в зерновки риса. Марганец — кофактор ферментов углеводного обмена. После поступления в растения он интенсивно транспортируется в побеги и используется в окислительно-восстановительных реакциях (39-41). В растениях выделен ряд Mn-стабилизирующих белков, вовлеченных в фотосинтез (40, 42, 43). Основная функция Se в растениях — участие в регуляции активности гликолитических и дыхательных ферментов (44-46). Наличие селена служит необходимым условием биосинтеза формиатдегидрогеназы — фермента, вызывающего окислительный распад муравьиной кислоты. Этот элемент активирует фумаразу (фумарат-гидратазу), катализирующую отщепление молекулы воды от яблочной кислоты в цикле Кребса, и нитратредуктазу, восстанавливающую нитрат до аммиака в процессе ассимиляции. Нуждается в селене и глутатионредуктаза — один из ключевых антиоксидантных ферментов (47, 48). В то же время высокие концентрации этого микроэлемента могут оказать негативное воздействие на метаболизм и ограничить вегетационный период растения (44, 49). Для растений риса критический уровень селена составляет 19 мкг/г сухой массы (50).
В течение 2009-2011 годов, когда проводились испытания потенциальных сеникантов, гидротермический режим различался. Температура воздуха в 2009 году в период посева и получения всходов риса (май—I декада июня) была на уровне средних многолетних значений, в течение вегетативного развития (июнь—начало августа) — превышала их на 1,0-4,3 °С, а в августе (налив зерновок) была на 0,6-1,3 °С ниже средней многолетней. В 2010 году температура воздуха в период вегетации риса аномально повысилась: начиная с III декады мая, она превышала средние многолетние значения на 3-6 °С. В I-II декадах мая в 2011 году выпало значительное количество осадков, а среднесуточные температуры воздуха были несколько ниже средних многолетних, из-за чего посев проводили позднее оптимальных сроков. В том же году в период всходов (май—I декада июня) температура воздуха была на 0,8-1,8 °С выше средней многолетней, в течение вегетативного роста превышала ее на 1,0-1,6 °С, а в III декаде августа—сентябре, когда шел налив зерновок, — на 0-1,0 °С. В целом температурный режим был благоприятным для роста, развития и формирования урожая риса (если не считать изменения сроков посева на более поздние).
Действие сеникантов и эффективность их применения прямо зависят от температурного режима, поэтому полученные результаты мы интерпретировали с учетом погодных условий года. Был установлен оптимальный срок сеникации (ранние сроки значительно сокращаются период налива зерновок, что отрицательно сказывается на их выполненности, поздние также не дают ожидаемого эффекта и лишь увеличивают затраты на выращивание культуры). Сравнили два срока обработки — через 10 сут после цветения главной метелки и в фазу молочно-восковой спелости зерна.
Эффективность сениканта оценивали по снижению влажности вегетативных органов и метелок с обязательным последующим учетом массы зерна с метелки и растения, массы 1000 зерен и урожайности. Эти показатели характеризуют физиологическое состояние растений в период созревания и конечный результат вызванных сеникацией изменений. В настоящей работе нами представлены результаты по одному среднепозднему сорту, однако следует учитывать, что у сортов с разным генотипом и тем более с неодинаковой продолжительностью вегетационного периода различается реакция на сениканты. Поэтому при адаптации технологии к иным сортам и погодным условиям требуются дополнительные исследования.
В 2009 году при сеникации посевов через 10 сут после завершения цветения главной метелки влажность метелок в 1-ю нед снижалась медленнее, чем при естественном созревания, что закономерно, так как основу сениканта составляет азотное удобрение. Из-за его воздействия на растения влажность метелок была на 0,3-2,0 % выше, чем в контроле (табл. 1). В течение 2-й нед влажность метелок активно снижалась (с сеникантами на основе мочевины она была на 2,1-4,2 % ниже, чем в контроле, аммонийной селитры — на 6,2-8,4 %. В последующие дни потеря влаги несколько замедлилась, но ее скорость все же оставалась выше при воздействии сеникантов на основе аммонийной селитры (особенно с добавлением Se и Mn). Через 30 сут после сеникации влажность метелок на этих растениях была на 2,3-2,8 % ниже, чем на контрольных.
Основное назначение сеникации — стимулировать аттракцию ассимилятов из вегетативных органов в зерновки (10). Уже через 1 нед после обработки растений сеникантами их сухая масса превышала контроль на 0,02-0,24 г, или на 1,4-17,0 %. На следующую неделю, когда наблюдалась наиболее интенсивная потеря влаги, при воздействии растворами мочевины и аммонийной селитры как в чистом виде, так и с добавлением марганца сухая масса метелок была на 2,7-5,8 % меньше, чем в контроле, а при добавлении селена — на 0,5-3,6 % больше. К концу вегетации только с сеникантами на основе мочевины масса метелок достоверно не отличалась от контроля, а при сеникации раствором аммонийной селитры (как в чистом виде, так и при добавлении микроэлементов) была даже на 7,0-9,7 % меньше. Это обусловлено влиянием сеникантов на скорость поступления ассимилятов в метелку: в 1-ю нед эта скорость была выше, чем в контроле, а затем постепенно снижалась.
Результатом изменения скорости созревания стало снижение урожайности: в наибольшей степени (на 14,3 и 12,7 %) — под влиянием сеникантов, содержащих Se. Негативные последствия от действия других сеникантов были не столь значительными, но тенденция обозначилась довольно четко. Следовательно, сеникацию непосредственно после цветения проводить нецелесообразно. Возможно, только в критической ситуации такая ранняя сеникация поможет спасти хотя бы некоторую часть урожая.
Проведение сеникации в начале фазы молочно-восковой спелости зерна (15 сут после цветения) выявило следующие закономерности. В период созревания зерновок влажность листьев и стеблей растений изменялась незначительно. Это обусловлено как биологией развития растений риса, у которых (в отличие от пшеницы и других зерновых культур) при созревании зерна листья и стебли продолжают функционировать, так и технологией возделывания. Произрастание риса на затопленном водой поле искажает показатели влажности стебля, так как на растении имеются листья и листовые влагалища, степень отмирания которых (и, как следствие, насыщенность водой) неодинаковы. Тем не менее можно констатировать, что по мере созревания растений различия по влажности вегетативных органов между контролем и вариантами с сеникацией увеличивались, хотя в целом не превышали 1-2 %. Добавление микроэлементов к раствору мочевины и аммонийной селитры вызывало большее снижение влажности стеблей, чем при их самостоятельном применении.
Препараты на основе мочевины сильнее подсушивали листья непосредственно после сеникации, на основе аммонийной селитры — действовали позднее. По достижении фазы полной спелости влажность листьев во всех вариантах была на 0,5-5,9 % ниже, чем в контроле, а добавленные микроэлементы усиливали потерю воды листьями.
На протяжении всех лет исследований достоверных различий по сухой массе листьев между контрольными и подвергнутыми сеникации растениями мы не отмечали, но наблюдалась общая тенденция к снижению этого показателя у последних. Масса стебля у обработанных растений была выше, чем в контроле.
Завершающая стадия созревания зерна — сложный биохимический процесс, связанный не только с поступлением ассимилятов в зерновку, но и с биосинтезом запасных веществ из низкомолекулярных соединений. При созревании происходит устойчивое снижение влажности на фоне увеличения массы сухого вещества и общего числа клеток как зародыша, так и эндосперма (51).
На ранних этапах созревания вода из семян удаляется метаболически, а затем физически за счет испарения (52, 53). Следовательно, снижение влажности зерновок служит легко учитываемым индикатором созревания (54). С биологической точки зрения созревшими принято считать семена, способные дать начало новому поколению. Техническая зрелость наступает несколько позднее. Условно наступление полной спелости фиксируют при влажности метелок 30 % от сырой массы.
При сеникации в фазу молочно-восковой спелости зерна снижение влажности метелок происходило неодинаково и определялось главным образом температурным режимом (рис.). За период от сеникации до сброса воды с рисового чека влажность метелок в 2009 году уменьшалась со средней скоростью 0,87 %/сут (0,79÷0,97 %), в 2010 году — 0,76 %/сут (0,73÷0,79 %), в 2011 году — 0,90 %/сут (0,85÷0,96 %), в контроле — соответственно 0,77; 0,71 и 0,80 %/сут. Этот процесс был наиболее интенсивным в 1-ю и 2-ю нед после сеникации — в среднем по вариантам 1,47 и 0,89 %/сут. Затем скорость уменьшения влажности метелок снижалась до 0,46-0,50 %/сут. В годы (2009 и 2011), когда погодные условия задерживали созревание, различия с контролем по скорости потери влаги метелками риса увеличивались.
Как показали наблюдения, влияние сеникантов на основе мочевины и аммонийной селитры несколько различается. В 1-ю нед после сеникации метелки быстрее теряли воду под воздействием препаратов на основе аммонийной селитры, на 2-ю нед — в вариантах на основе мочевины; затем до полной спелости скорость потери воды приблизительно сравнялась. К сбросу воды с рисового поля влажность метелок на растениях, обработанных сеникантами на основе аммонийной селитры, оказалась ниже, чем при использовании мочевины. Это обусловлено неодинаковой скоростью проникновения препаратов в лист вследствие особенностей механизма поглощения амидного и аммонийного азота. Аммонийная селитра быстрее попадает в растение, и ее действие начинается несколько раньше. На эффективности сениканта также сказывается бóльшая затрата энергии на вовлечение аммонийного азота в метаболизм. Включение в состав сениканта Mn и Se усиливало действие аммонийной селитры и мочевины, причем в большей степени при применении селена, что связано с его влиянием на водный режим растения. Отметим, что увеличение концентрации селена в растворе ускоряет потерю влаги метелками и, следовательно, позволяет раньше приступить к уборке (данные не приведены).
При температуре воздуха в период налива зерновок, близкой к средним многолетним значениям (2009 и 2011 годы), метелки на растениях, подвергнутых сеникации, достигали 30 % влажности на 1-6 сут раньше, чем в контроле. Впоследствии они теряли влагу быстрее, и перед сбросом воды их влажность была на 0,69-7,39 % ниже, чем в контроле. Если температура воздуха в этот период была значительно выше средних многолетних значений (2010 год), влажность метелок на обработанных растениях снижалась до 30 % на 4-10 сут раньше, но в дальнейшем у контрольных растений скорость этого процесса возрастала, различия сглаживались и к сбросу воды с чека составляли лишь 0,66-2,78 %.
В процессе созревания масса метелок достигает максимума к фазе полной спелости зерна. Сениканты стимулировали накопление сухого вещества в зерновках. В 1-ю нед после обработки его количество увеличивалось со скоростью 0,072-0,082 г/(растение · сут), что на 24,1-41,4 % выше, чем у контрольных растений. В дальнейшем (до завершения налива зерновок) интенсивность накопления сухого вещества имела тенденцию к постепенному снижению. В контроле накопление сухого вещества в метелках усиливалось в последнюю неделю перед завершением налива зерновок (табл. 2).
На протяжении первых 2 нед фазы молочно-восковой спелости зерна увеличение сухой массы метелок происходило быстрее, затем процесс несколько замедлялся, а к завершению созревания вновь ускорялся, особенно в контроле. При сеникации сухая масса метелок за 1-ю нед увеличивалась на 35,4 %, за 2-ю — на 20,3 %, за 3-ю — на 10,6 %, 4-ю — на 12,2 %. За то же время у контрольных растений этот показатель вырос соответственно лишь на 27,4; 14,8; 10,8 и 18,4 % (см. рис.).
В 2009 году, который мы условно обозначили как холодный (с понижением температуры в период налива зерновок), отличия от контроля по сухой массе метелок обнаруживались через 1 нед после сеникации и оценивались в 5,3-6,4 % при использовании мочевины (как отдельно, так и с добавлением Mn и Se) и 3,7-4,3 % — в вариантах с сеникантами на основе аммонийной селитры. За следующую неделю разница увеличилась до 5,9-11,4 % (препараты с мочевиной) и 9,1-11,9 % (с аммонийной селитрой), а в последнюю неделю налива зерновок приток ассимилятов у контрольных растений происходил в 1,3-1,6 раза быстрее, что обусловило уменьшение различий на 1,0-5,8 %. В 2010 году в начале налива зерновок дневные температуры поднимались до 40 °С. В результате сочетания эффектов температур и сеникантов различия между растениями по сухой массе метелок были значительно меньше, чем в 2009 году: в 1-ю нед — 0,7-3,6 %, на 2-ю — 5,3-8,6 %, на 3-ю — несущественными, но далее на растениях, подвергнутых сеникации, ассимиляты стали интенсивно поступать в зерновки, и различия с контролем возросли до 4,5-13,9 %. В 2011 году (по температурному режиму в период налива зерновок близок к 2009 году) при идентичных закономерностях накопления сухого вещества метелками отмечали более существенное положительное влияние сеникантов. Скорость увеличения сухой массы метелок на обработанных растениях в 1-ю и 2-ю нед была на 38,2-79,4 % выше, чем в контроле, с 14-х по 21-е сут после сеникации мочевиной и аммонийной селитрой оставалась соответственно на 58,6 и 62,1 % выше, чем в контроле, при модификации марганцем — лишь на 6,9-17,2 %, а в присутствии селена в составе сеникантов уменьшилась соответственно на 17,2 и 31,0 %. В течение 4-й нед ассимиляты в метелку поступали в 1,3-3,1 раза медленнее, чем в контроле. Вследствие такой динамики накопления сухого вещества сухая масса метелок у обработанных растений была больше, чем в контроле: через 1 нед — на 9,3-13,0 %, через 2 нед — на 14,0-21,5 %, а при достижении полной спелости — на 2,9-13,0 %. Иными словами, если бы возникла необходимость начать уборку в более ранние сроки, сеникация позволила бы лучше сохранить урожай.
Полученные нами результаты свидетельствуют, что в конечном итоге мочевина и аммонийная селитра существенно не различаются по влиянию на созревание зерновок у риса, несмотря на неодинаковую динамику влажности и сухой массы метелок. Сеникация посевов водными растворами мочевины и аммонийной селитры, модифицированными марганцем, обеспечивает лучший налив зерновок. Включение селена в состав сениканта ускоряет снижение влажности метелок, что ведет к замедлению скорости накопления сухого вещества в конце фазы молочно-восковой спелости.
Интенсивный отток ассимилятов из вегетативных органов растений риса в метелку под воздействием сеникантов — безусловно, положительный эффект этого агроприема, как и более быстрое или значительное снижение ее влажности, что позволяет уменьшить затраты на уборку и послеуборочные мероприятия. Однако при благоприятных условиях, которые могут наступить после сеникации, сокращение периода налива зерновок может обернуться потерей урожая. Поэтому при выборе сеникантов следует отдавать предпочтение тем, которые при благоприятном температурном режиме не вызывают снижения урожая по сравнению с посевами без сеникации. В холодном 2009 году мы отмечали незначительное уменьшение урожайности при сеникции раствором мочевины с добавлением Se (табл. 3), тогда как препараты на основе аммонийной селитры показали лучший результат: урожайность повышалась, причем в варианте аммонийная селитра + Mn — существенно. Рост урожайности был обусловлен увеличением массы зерна с метелки и массы 1000 зерен. В 2010 году аномально высокие температуры воздуха в период налива зерновок оказались критическими для величины урожая. Сеникация растворами мочевины (в чистом виде и с добавлением Mn) не купировала эти негативные последствия, и урожайность риса в этих вариантах не отличалась от контроля. Обработка раствором мочевины с добавлением Se снизила урожайность на 12,2 % (главным образом вследствие уменьшения массы 1000 зерен). Сениканты на основе аммонийной селитры, напротив, способствовали формированию урожая, который был на 7,9-14,7 % выше, чем в контроле. Менее всего его росту благоприятствовала сеникация раствором аммонийной селитры с добавлением Se. В 2011 году температура ниже средних многолетних значений и осадки в мае задержали посев, но созревание шло при температуре воздуха, близкой к средней многолетней (отклонение 0-1,0 °С). В этот год сеникация раствором мочевины и аммонийной селитры существенно не повлияла на урожайность, но при использовании растворов, модифицированных марганцем, прибавка урожая относительно контроля была статистически значимой (НСР05 = 0,38 т/га) и составила 7,9 и 9,3 %, а в присутствии Se обозначилась тенденция к ее снижению. В то же время обращает на себя внимание факт снижения влажности зерна к уборке урожая под действием сеникантов. Поэтому целесообразность сеникации обусловлена как минимум тем, что затраты на ее проведение не выше стоимости мероприятий по снижению влажности зерна до стандартной.
Таким образом, сеникация — важнейший прием целенаправленного регулирования метаболизма на завершающих стадиях развития растений риса, особенно в условиях зоны рискованного возделывания этой культуры. И хотя в настоящее время сеникация посевов риса используется мало (55, 56), что обусловлено главным образом продолжительным теплым периодом в многолетнем цикле колебаний температур (57), высока вероятность наступления в ближайшие годы периода с неблагоприятными для созревания и уборки риса погодными условиями, и востребованность этого приема возрастет. Отдельного внимания заслуживает универсальность предложенной агротехнологии, позволяющей при необходимости оперативно корректировать сроки уборки у сортов любой группы спелости. Для результативного применения этого метода на современных сортах риса следует предварительно изучить у них процесс созревания зерновок при сеникации. Также необходим поиск новых сеникантов, безопасных для человека и окружающей среды (39). Актуальны исследования по уточнению срока сеникации (40). По нашим данным, при этом следует руководствоваться суммой эффективных температур (для риса — более 15 °С) (40).
Итак, показано, что эффективность сеникации на посевах среднепоздних сортов риса зависит от состава сениканта, срока обработки и погодных условий. Сеникацию следует применять при запаздывании созревания, наблюдаемом чаще всего на поздних, изреженных или получивших избыточное азотное питание посевах. Наиболее подходящие сениканты — водные растворы аммонийной селитры (15 кг/га) с добавлением марганца (400 г/га) или селена (200 г/га). Модифицированный селеном раствор следует использовать при необходимости приступить к уборке через 14-20 сут, при этом увеличение концентрации селена в растворе ускоряет потерю влаги метелками и сокращает срок до начала уборки. Обработка растений раствором, содержащим марганец, стимулирует синтез ассимилятов и их аттракцию в зерновку, что сопровождается не столь быстрым завершением онтогенеза, как при модификации селеном, но существенным ростом урожая вследствие увеличения индивидуальной продуктивности растений. Назначать сеникацию посевов риса непосредственно по окончанию фазы цветения нецелесообразно, так как быстрое завершение вегетации снижает урожай. Оптимальным сроком сеникации следует считать начало фазы молочно-восковой спелости. В годы, когда температура воздуха ниже и/или близка к средней многолетней, сеникация не только быстрее (на 5-6 сут) и значительнее (на 0,69-7,39 %) снижает влажность метелок, но и повышает урожайность зерна на 4,2-9,3 %. В годы с благоприятными для созревания риса погодными условиями сеникация обеспечивает снижение влажности метелок к сбросу воды с рисового чека лишь на 0,66-2,78 %, но увеличивает урожайность на 7,9-14,7 % благодаря дополнительному поступлению азота в растения (при достаточной для использования этого азота продолжительности периода благоприятных температур такая подкормка способствует повышению продуктивности побегов 2-3-го порядка).
1. Динамика созревания и урожайность риса (Oryza sativa L.) сорта Хазар после сеникации посевов в фазу молочной |
||||||||||
Вариант (сеникант) |
Влажность, % |
Сухая масса, г/растение |
Скорость накопления сухого |
Урожайность, т/га |
||||||
6-е сут |
12-е сут |
30-е сут |
6-е сут |
12-е сут |
30-е сут |
0-6-е сут |
7-12-е сут |
13-30-е сут |
||
Контроль (без сеникации) |
40,6±0,81 |
40,4±1,01 |
19,8±0,46 |
1,42±0,10 |
2,24±0,11 |
2,59±0,16 |
0,052±0,018 |
0,117±0,005 |
0,018±0,001 |
12,6±0,4 |
Мочевина |
42,6±0,85 |
38,3±0,96 |
22,3±0,52 |
1,44±0,06 |
2,30±0,12 |
2,61±0,15 |
0,055±0,019 |
0,123±0,005 |
0,016±0,002 |
12,5±0,5 |
Мочевина + Mn |
42,4±0,84 |
37,3±0,93 |
20,7±0,49 |
1,48±0,07 |
2,37±0,12 |
2,64±0,12 |
0,062±0,022 |
0,127±0,005 |
0,014±0,001 |
12,8±0,3 |
Мочевина + Se |
41,1±0,82 |
36,2±0,91 |
19,0±0,44 |
1,66±0,09 |
2,23±0,13 |
2,48±0,11 |
0,092±0,033 |
0,081±0,003 |
0,013±0,003 |
10,8±0,2 |
Аммонийная селитра |
42,8±0,86 |
34,2±0,86 |
17,5±0,41 |
1,55±0,07 |
2,33±0,14 |
2,40±0,15 |
0,073±0,028 |
0,111±0,005 |
0,004±0,0002 |
12,3±0,4 |
Аммонийная селитра + Mn |
41,2±0,82 |
33,8±0,85 |
17,2±0,40 |
1,54±0,06 |
2,35±0,12 |
2,41±0,11 |
0,072±0,025 |
0,116±0,005 |
0,003±0,0004 |
12,7±0,3 |
Аммонийная селитра + Se |
38,3±0,77 |
32,0±0,80 |
17,0±0,39 |
1,58±0,07 |
2,16±0,11 |
2,34±0,12 |
0,078±0,027 |
0,083±0,003 |
0,009±0,0005 |
11,0±0,4 |
НСР05 |
|
|
|
0,11 |
0,14 |
0,17 |
|
|
|
0,40 |
2. Скорость накопления сухого вещества в метелках риса (Oryza sativa L.) сорта Хазар после сеникации посевов в фазу молочно-восковой спелости зерна, г/(растение · сут) (M±SEM, Краснодарский край) |
|||||||||||||
Вариант |
2009 год |
2010 год |
2011 год |
||||||||||
0-5-е сут |
6-12-е сут |
13-19-е сут |
20-28-е сут |
0-7-е сут |
8-14-е сут |
15-21-е сут |
22-28-е сут |
29-35-е сут |
0-7-е сут |
8-14-е сут |
15-21-е сут |
22-28-е сут |
|
1 |
0,058±0,003 |
0,046±0,002 |
0,027±0,002 |
0,057±0,003 |
0,054±0,003 |
0,020±0,001 |
0,033±0,002 |
0,021±0,001 |
0,016±0,001 |
0,057±0,002 |
0,034±0,002 |
0,029±0,002 |
0,059±0,002 |
2 |
0,080±0,005 |
0,049±0,002 |
0,043±0,003 |
0,040±0,002 |
0,056±0,003 |
0,034±0,002 |
0,019±0,001 |
0,046±0,002 |
0,029±0,002 |
0,084±0,003 |
0,047±0,003 |
0,046±0,003 |
0,020±0,001 |
3 |
0,078±0,005 |
0,067±0,003 |
0,044±0,003 |
0,028±0,002 |
0,063±0,003 |
0,030±0,002 |
0,023±0,001 |
0,053±0,002 |
0,016±0,001 |
0,094±0,001 |
0,054±0,003 |
0,031±0,002 |
0,019±0,001 |
4 |
0,082±0,005 |
0,061±0,003 |
0,039±0,002 |
0,029±0,002 |
0,057±0,003 |
0,029±0,002 |
0,026±0,001 |
0,040±0,002 |
0,006±0,000 |
0,079±0,003 |
0,061±0,003 |
0,024±0,001 |
0,024±0,001 |
5 |
0,072±0,004 |
0,064±0,003 |
0,039±0,002 |
0,023±0,001 |
0,047±0,002 |
0,041±0,002 |
0,016±0,001 |
0,043±0,002 |
0,027±0,002 |
0,081±0,003 |
0,047±0,003 |
0,047±0,003 |
0,026±0,001 |
6 |
0,074±0,004 |
0,071±0,004 |
0,034±0,002 |
0,043±0,003 |
0,061±0,003 |
0,030±0,002 |
0,017±0,001 |
0,057±0,003 |
0,016±0,001 |
0,087±0,003 |
0,056±0,003 |
0,034±0,002 |
0,047±0,002 |
7 |
0,072±0,004 |
0,069±0,003 |
0,037±0,002 |
0,032±0,002 |
0,050±0,003 |
0,036±0,002 |
0,014±0,001 |
0,043±0,002 |
0,026±0,002 |
0,083±0,003 |
0,056±0,003 |
0,020±0,001 |
0,019±0,001 |
НСР05 |
0,006 |
0,003 |
0,004 |
0,004 |
0,003 |
0,002 |
0,001 |
0,001 |
0,002 |
0,002 |
0,003 |
0,003 |
0,001 |
Примечание. 1 — контроль (без сеникации), 2 — мочевина, 3 — мочевина + Mn, 4 — мочевина + Se, 5 — аммонийная селитра, 6 — аммонийная селитра + Mn, |
|||||||||||||
3. Урожайность и структура урожая риса (Oryza sativa L.) сорта Хазар после сеникации посевов в фазу молочно-восковой спелости зерна по годам ис-следований (M±SEM, Краснодарский край) |
||||||||||||
Вариант |
2009 год |
2010 год |
2011 год |
|||||||||
У, т/га |
МЗМ, г |
ПЗ, % |
М1000, г |
У, т/га |
МЗМ, г |
ПЗ, % |
М1000, г |
У, т/га |
МЗМ, г |
ПЗ, % |
М1000, г |
|
1 |
11,44±0,34 |
3,19±0,18 |
0,9±0,1 |
27,88±1,39 |
4,82±0,29 |
2,06±0,12 |
6,3±0,4 |
26,95±0,81 |
8,62±0,34 |
2,41±0,13 |
19,9±1,0 |
28,85±1,15 |
2 |
11,49±0,34 |
3,24±0,18 |
1,8±0,1 |
29,85±1,49 |
4,84±0,29 |
2,14±0,13 |
5,2±0,4 |
27,08±0,81 |
8,90±0,36 |
2,56±0,14 |
18,8±1,1 |
28,89±1,16 |
3 |
11,66±0,35 |
3,29±0,19 |
1,7±0,1 |
29,98±1,50 |
4,86±0,29 |
2,17±0,13 |
5,2±0,4 |
27,20±0,80 |
9,30±0,37 |
2,62±0,14 |
17,3±1,0 |
29,20±1,17 |
4 |
11,08±0,33 |
3,21±0,18 |
1,6±0,1 |
28,98±1,45 |
4,23±0,25 |
2,08±0,12 |
5,6±0,4 |
26,02±0,78 |
8,36±0,33 |
2,55±0,14 |
16,7±1,0 |
29,01±1,16 |
5 |
11,58±0,35 |
3,20±0,18 |
1,6±0,1 |
29,12±1,46 |
5,40±0,32 |
2,42±0,15 |
5,2±0,4 |
27,10±0,81 |
8,94±0,36 |
2,61±0,14 |
19,4±1,2 |
29,03±1,16 |
6 |
11,92±0,36 |
3,39±0,19 |
1,6±0,1 |
29,25±1,46 |
5,53±0,33 |
2,43±0,15 |
5,9±0,4 |
27,15±0,81 |
9,42±0,38 |
2,81±0,15 |
17,5±1,1 |
29,37±1,17 |
7 |
11,61±0,35 |
3,09±0,18 |
1,6±0,1 |
29,00±1,45 |
5,20±0,31 |
2,30±0,14 |
5,2±0,4 |
27,00±0,81 |
8,54±0,34 |
2,41±0,13 |
18,9±1,1 |
28,401,14 |
НСР05 |
0,35 |
0,19 |
|
1,49 |
0,38 |
0,27 |
|
0,74 |
0,38 |
0,2 |
|
1,12 |
Примечание. 1 — контроль (без сеникации), 2 — мочевина, 3 — мочевина + Mn, 4 — мочевина + Se, 5 — аммонийная селитра, |
||||||||||||
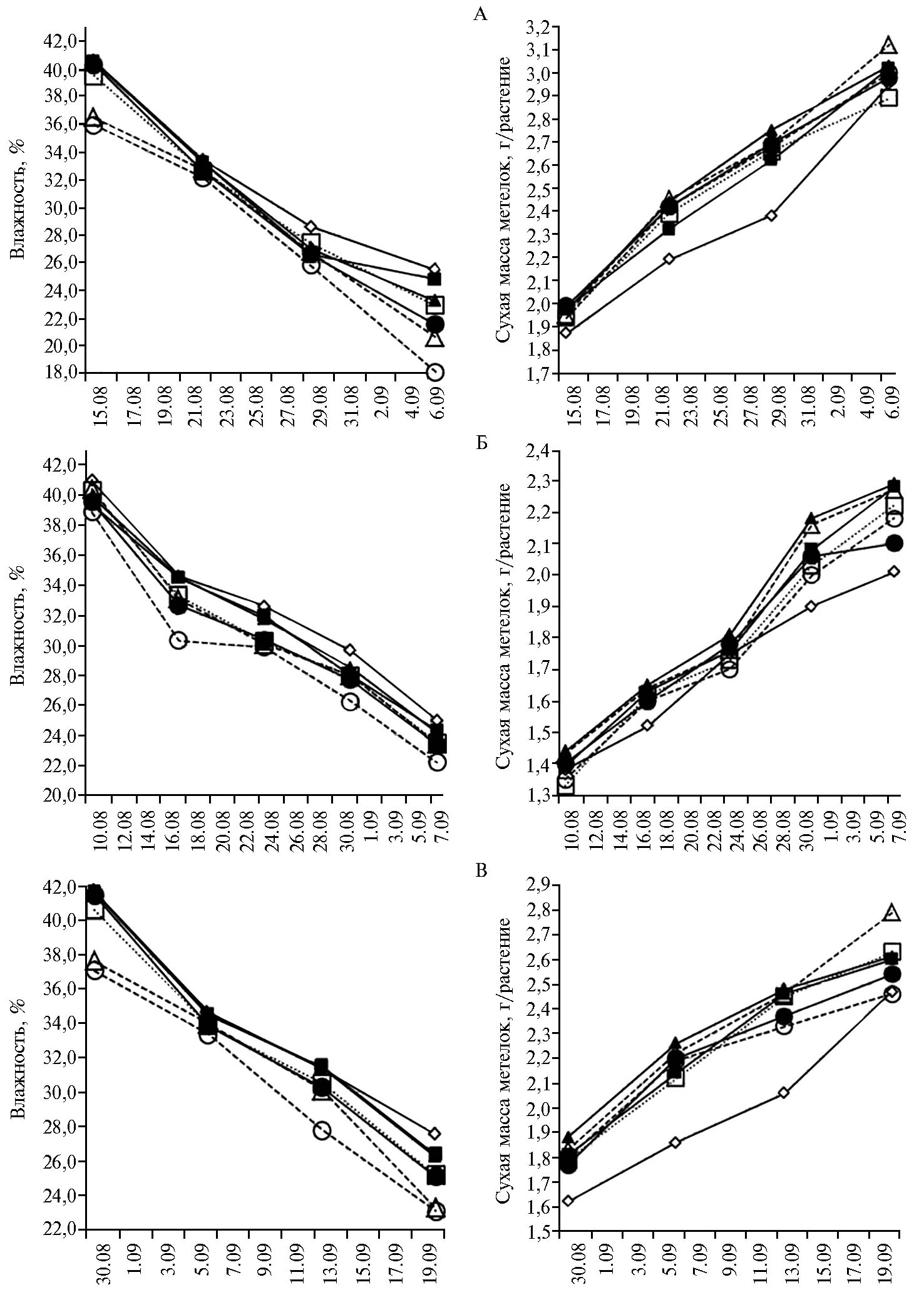
Динамика влажности (слева) и сухой массы (справа) метелок у риса (Oryza sativa L.) сорта Хазар после сеникации посевов в фазу молочно-восковой спелости зерна в 2009 (А), 2010 (Б) и 2011 (В) годах:  — контроль (без сеникации),
— контроль (без сеникации),  — мочевина,
— мочевина,  — мочевина + Mn,
— мочевина + Mn,  — мочевина + Se,
— мочевина + Se,  — аммонийная селитра,
— аммонийная селитра,  — аммонийная селитра + Mn,
— аммонийная селитра + Mn,  — аммонийная селитра + Se (Краснодарский край).
— аммонийная селитра + Se (Краснодарский край).
ЛИТЕРАТУРА
- Paddy rice production worldwide 2017-2018, by country. Режим доступа: https://www.sta-tista.com/statistics/255937/leading-rice-producers-worldwide/. Дата обращения 02.06.2019.
- Chauhan B.S, Jabran K., Mahajan G. Rice production worldwide. Springer International Publising AG, Sqitzerland, 2017 (doi: 10.1007/978-3-319-47516-5).
- Kraehmer H., Thomas C., Vidotto F. Rice production in Europe. In: Rice production worldwide /B. Chauhan, K. Jabran, G. Mahajan (eds.). Springer, Cham, 2017: 93-116 (doi: 10.1007/978-3-319-47516-5_4).
- Tesio F., Tabacchi M., Cerioli S., Follis F. Sustainable hybrid rice cultivation in Italy. A review. Agron. Sustain. Dev., 2014, 34: 93-102.
- Beser N., Surek H., Sahin M. Search of effective resistant genes to the rice blast pathogen (Magnaporthe grisea) under field conditions in Turkey. Fresenius Environmental Bulletin, 2015, 24: 791-795.
- Воробьев Н.В. Физиологические основы формирования урожая риса. Краснодар, 2013.
- de Vries G.E. Growing rice in Canada. Trends in Plant Science, 2000, 5(1): 8 (doi: 10.1016/S1360-1385(99)01535-6).
- Clement C., Burrus M., Audran J.-C. Floral organ growth and carbohydrate content during pollen development in Lilium. American Journal of Botany, 1996, 83(4): 459-469 (doi: 10.2307/2446215).
- Lemoine R., La Camera S., Atanassova R., Dédaldéchamp F., Allario T., Pourtau N., Bonnemain J.-L., Laloi M., Coutos-Thévenot P., Maurousset L., Faucher M., Girousse C., Lemonnier P., Parrilla J., Durand M. Source-to-sink transport of sugar and regulation by environmental factors. Front. Plant Sci., 2013, 4: 272 (doi: 10.3389/fpls.2013.00272).
- Ray S., Choudhuri M.A. Mobilization of metabolites from leaves to grains as the cause of monocarpic senescence in rice. PlantPhysiol., 1981, 68: 1345-1348.
- Lim P.O., Kim H.J., Nam H.G. Leaf senescence. Annual Review of Plant Biology, 2007, 58: 115-136 (doi: 10.1146/annurev.arplant.57.032905.105316).
- Li Z., Pan X., Guo X., Fan K., Lin W. Physiological and transcriptome analyses of early leaf senescence for ospls1 mutant rice (Oryza sativa L.) during the grain-filling stage. Int. J. Mol. Sci., 2019, 20: 1098 (doi: 10.3390/ijms20051098).
- Букреева Г.И., Грицай Т.И., Домченко М.И. Особенности формирования качества зерна озимой мягкой пшеницы в контрастных погодных условиях. Труды Кубанского государственного аграрного университета, 2015, 56: 85-91.
- Евдокимов М.Г., Юсов В.С., Татина Б.М., Андреева В.В. Формирование и налив зерна яровой твердой пшеницы в условиях лесостепи Западной Сибири. Вестник Алтайского государственного аграрного университета, 2015, 11(133): 5-9.
- Wojciechowska N., Sobieszczuk-Nowicka E., Bagniewska‐Zadworna A. Plant organ senescence — regulation by manifold pathways. Plant Biol. J., 2018, 20: 167-181 (doi: 10.1111/plb.12672).
- Афонин Н.М. Эффективность десикации и сеникации посевов кукурузы при выращивании на зерно. Кукуруза и сорго, 2010, 3: 14-16.
- Колосова Е.Н. Влияние сеникации на продуктивность гречихи. Мат. Межд. науч.-практ. конф. «Научное обеспечение агропромышленного производства». Курск, 2010: 214-216.
- Distelfeld A., Avni R., Fischer A.M. Senescence, nutrient remobilization, and yield in wheat and barley. J. Exp. Bot., 2014, 65: 3783-3798 (doi: 10.1093/jxb/ert477).
- Jibran R., Hunter A., Dijkwel P. Hormonal regulation of leaf senescence through integration of developmental and stress signals. Plant Mol. Biol., 2013, 82: 547-561 (doi: 10.1007/s11103-013-0043-2).
- Lin M., Pang C., Fan S., Song M., Wei H., Yu S. Global analysis of the Gossypium hirsutum L. transcriptome during leaf senescence by RNA-seq. BMC Plant Biology, 2015, 15: 43 (doi: 10.1186/s12870-015-0433-5).
- Кушу А.А. Повышения продуктивности семеноводческих рисовых полей путем использования сеникации в условиях Адыгеи. Автореф. канд. дис. Краснодар, 1995.
- Воробьев П.Н. Сеникация на семенниках капусты белокочанной. В сб.: Селекция, семеноводство и технология возделывания овощных культур. Воронеж, 1989: 77-83.
- Молявко А.А., Марухленко А.В., Еренкова Л.А., Борисова Н.П. Влияние сеникации и других приемов на урожайность картофеля. Защита и карантин растений, 2017, 11: 30-31.
- Goswamy B.K., Srivastava G.C. Effect of foliar application of urea on leaf senescence and photosynthesis in sunflower (Helianthus annuus L.). Photosynthetica, 1988, 22(1): 99-104.
- Романько Ю.А., Мельник А.В. Урожайность и качество семян сои при применении сеникации и десикации в условиях левобережной лесостепи Украины. Известия Самарской государственной сельскохозяйственной академии, 2016, 1(2): 15-18.
- Серен К.Д., Игнатьев Л.А. Сеникация посевов сои (Glicine hispida Maxim.) в сухостепной зоне Республики Тыва. Агрохимия, 2008, 2: 50-56.
- Павлютина И.П., Моисеенко И.Я., Лихачев Б.С. Приемы ускорения созревания семян сои. Кормопроизводство, 2005, 1: 24-27.
- Шеуджен А.Х., Бондарева Т.Н. Агрохимия. Ч. 2. Методика агрохимических исследований. Краснодар, 2015.
- Алешин Е.П., Авакян К.М., Подлесный И.В., Семенова Л.М., Коровянский А.Г., Рыбаченко С.И. Сеникация риса жидкими комплексными удобрениями. Бюллетень НТИ ВНИИ риса, 1986, 36: 37-40.
- Craft J.C., Lindsey L.E., Barker D.J., Lindsey A.J. Quantification of soybean leaf senescence and maturation as impacted by soil- and foliar-applied nitrogen. Crop, Forage & Turfgrass Management, 2019, 5: 180051 (doi: 10.2134/cftm2018.07.0051).
- Springer A., Kang C.H., Rustgi S., von Wettstein D., Reinbothe C., Pollmann S., Reinbothe S. Programmed chloroplast destruction during leaf senescence involves 13-lipoxygenase (13-LOX). PNAS USA, 2016, 113(12): 3383-3388 (doi: 10.1073/pnas.1525747113).
- Panda D., Sarkar R.K. Natural leaf senescence: Probed by chlorophyll fluorescence, CO2 photosynthetic rate and antioxidant enzyme activities during grain filling in different rice cultivars. Physiol. Mol. Biol. Plants, 2013, 19(1): 43-51 (doi: 10.1007/s12298-012-0142-6).
- Avila-Ospina L., Marmagne A., Talbotec J., Krupinska K., Masclaux-Daubresse C. The identification of new cytosolic glutamine synthetase and asparagine synthetase genes in barley (Hordeum vulgare L.), and their expression during leaf senescence. J. Exp. Bot., 2015, 66(7): 2013-2026 (doi: 10.1093/jxb/erv003).
- Britto D.T., Kronzucker H.J. NH4+ toxicity in higher plants: a critical review. Journal of Plant Physiology, 2002, 159(6): 567-584 (doi: 10.1078/0176-1617-0774).
- Bieker S., Zentgraf U. Plant senescence and nitrogen mobilization and signaling. In: Senescence and senescence-related disorders /Z. Wang, H. Inuzuka (eds.). Intech Open, 2013: 53-83 (doi: 10.5772/54392).
- Nawaz F., Naeem M., Ashraf M.Y., Tahir M.N., Zulfiqar B., Salahuddin M., Shabbir R.N., Aslam M. Selenium supplementation affects physiological and biochemical processes to improve fodder yield and quality of maize (Zea mays L.) under water deficit conditions. Front. Plant Sci., 2016, 7: 1438 (doi: 10.3389/fpls.2016.01438).
- Takano J., Wada M., Ludewig U., Schaaf G., von Wirén N., Fujiwara T. The Arabidopsis major intrinsic protein NIP5;1 is essential for efficient boron uptake and plant development under boron limitation. Plant Cell, 2006, 18: 1498-1509 (doi: 10.1105/tpc.106.041640).
- Socha A.L., Guerinot M.L. Mn-euvering manganese: The role of transporter gene family members in manganese uptake and mobilization in plants. Front. Plant Sci., 2014, 5: 106 (doi: 10.3389/fpls.2014.00106).
- Шеуджен А.Х. Теория и практика применения микро- и ультрамикроудобрений в рисоводстве. Майкоп, 2016.
- Sheudzhen A.Kh., Bondareva T.N., Kharchenko P.N., Doroshev I.A., Khurum Kh.D. Time determination of conducting senication of rice crops taking into account the sum of effective temperatures. Russian Agricultural Sciences, 2018, 44(6): 499-504 (doi: 10.3103/S1068367418060149).
- Page V., Weisskopf L., Feller U. Heavy metals in white lupine: uptake. Root-to-shoot transfer and redistribution within the plant. New Phytologist, 2006, 171: 329-241 (doi: 10.1111/j.1469-8137.2006.01756.x).
- Wyman A. J., Yocum C. F. Structure and activity of the photosystem II manganese-stabilizing protein: role of the conserved disulfide bond. Photosynthesis Research, 2005, 85: 359-372 (doi: 10.1007/s11120-005-7385-9).
- Popelkova H., Yocumb C.F. PsbO, the manganese-stabilizing protein: analysis of the structure—function relations that provide insights into its role in photosystem II. Journal of Photochemistry and Photobiology B: Biology, 2011, 104(1-2): 179-190 (doi: 10.1016/j.jphotobiol.2011.01.015).
- Шеуджен А.Х., Бондарева Т.Н., Дорошев И.А., Есипенко С.В. Селеновые удобрения на посевах риса. Майкоп, 2017.
- Seppanen M., Turakainen M., Hartikainen H. Selenium effects on oxidative stress in potato. Plant Science, 2003, 165: 311-319 (doi: 10.1016/S0168-9452(03)00085-2).
- Irmak S. Effects of selenium application on plant growth and some quality parameters in peanut (Arachis hypogaea). Pak. J. Biol. Sci., 2017, 20(2): 92-99 (doi: 10.3923/pjbs.2017.92.99).
- Couto N., Wood J., Barber J. The role of glutathione reductase and related enzymes on cellular redox homoeostasis network. Free Radic. Biol. Med., 2016, 95: 27-42 (doi: 10.1016/j.freeradbiomed.2016.02.028).
- Trivedi D.K., Gill S.S., Yadav S., Tuteja N. Genome-wide analysis of glutathione reductase (GR) genes from rice and Arabidopsis. Plant Signal. Behav., 2013, 8(2): e23021 (doi: 10.4161/psb.23021).
- Garousi F. Toxicity of selenium, application of selenium in fertilizers, selenium treatment of seeds, and selenium in edible parts of plants. Acta Universitatis Sapientiae, Alimentaria, 2017, 10: 61-74 (doi: 10.1515/ausal-2017-0004).
- Rani N., Dhillon К.S., Dhillon S.K. Critical levels of selenium in different crops grown in an alkaline silty loam soil treated with selenite-Se. Plant Soil, 2005, 277: 367-374 (doi: 10.1007/s11104-005-8161-5).
- Wu X., Li J., Li D., Liu C.M. Rice caryopsis development II: Dynamic changes in the endosperm. J. Integr. Plant Biol., 2016, 58: 786-798 (doi: 10.1111/jipb.12488).
- Brooking I.R. Maize ear moisture during grain-filling, and its relation to physiological maturity and grain-drying. Field Crops Research, 1990, 23(1): 55-68 (doi: 10.1016/0378-4290(90)90097-U).
- Maiorano А., Fanchini D., Donatellib M. MIMYCS.Moisture, a process-based model of moisture content in developing maize kernels. European Journal of Agronomy, 2014, 59: 86-95 (doi: 10.1016/j.eja.2014.05.011).
- Lin L., He Y., Xiao Z., Zhao K., Dong T., Nie P. Rapid-detection sensor for rice grain moisture based on NIR spectroscopy. Appl. Sci., 2019, 9(8): 1654 (doi: 10.3390/app9081654).
- Молоков Л.Г., Похно С.Л., Зеленева И.А., Григорьев В.А., Дворник Е.Д. Сеникация посевов риса: за и против. Рисоводство, 2013, 23: 68-72.
- Шхапацев А.К. Влияние микроэлементов, регуляторов роста и сеникации посевов на эффективность азотных удобрений и урожайность риса. Новые технологии, 2006, 2: 44-46.
- Шеуджен А.Х., Галкин Г.А., Бондарева Т.Н. Теплообеспеченность периода вегетации и урожайность риса. Рисоводство, 2007, 11: 24-28.