








УДК 636.2:591.133.2:636.084
ОБ ОСОБЕННОСТЯХ ОБМЕНА АЗОТИСТЫХ СОЕДИНЕНИЙ КОРМА В РУБЦЕ БЫКОВ В ЗАВИСИМОСТИ ОТ ПЕРИОДА ПОСЛЕ КОРМЛЕНИЯ
Н.С. ШЕВЕЛЕВ, Н.А. ПОЛОЗКОВА
У быков черно-пестрой породы с фистулами рубца через 3, 6 и 10 ч после кормления в цельном и разделенном на фракции содержимом рубца, а также в рубцовой жидкости определяли содержание азотистых соединений. Показано, что вследствие гомеостатических процессов в течение всего периода наблюдения в рубцовой жидкости поддерживается сравнительно постоянное количественное соотношение азотистых соединений.
Ключевые слова: крупный рогатый скот, фистулированные животные, жвачные, рубец, фракции содержимого рубца, азотистые соединения, гомеостаз.
При изучении обменных процессов в пищеварительном тракте жвачных животных, которое необходимо для разработки сбалансированных рационов, азотистые соединения вызывают наибольший интерес в связи с накоплением данных о динамике процессов рубцового метаболизма и микробиологии рубца. В последнее десятилетие установлено, что химус представляет собой структурированное образование. В его формировании и гомеостазировании важную роль выполняет плотная эндогенная фракция (ПЭФ) (1, 2), основу которой составляет слизь, содержащая мукополисахариды (3 %), белки (2 %) и соли (до 1 %) (3). Установлено, что ПЭФ может участвовать в обмене минеральных веществ в пищеварительном тракте и влиять на их всасывание, а также упорядочивает взаимодействие гидролитических ферментов с пищевыми частицами в процессе полостного пищеварения (4, 5).
Целью нашей работы было изучение динамики распределения азотистых соединений между фракциями рубцового содержимого в зависимости от периода времени после кормления животных.
Методика. Исследования выполняли на зоостанции Российского государственного аграрного университета (РГАУ-МСХА им. К.А. Тимирязева) на четырех 12-16-месячных быках черно-пестрой породы (средняя живая масса — 250-300 кг) с фистулами рубца, наложенными по методу В.А. Басова (4). В течение опыта животные получали сено-концентратный рацион, составленный согласно детализированным нормам кормления (6). Общее физическое состояние быков в период эксперимента было в пределах физиологической нормы.
Опыт проводили методом периодов. Через фистулы спустя 3, 6 и 10 ч после кормления отбирали пробы рубцового содержимого (последнюю — практически натощак). Для получения рубцовой жидкости (РЖ) усредненную пробу цельного рубцового содержимого (ЦРС) от всех животных отжимали через четыре слоя марли. ЦРС разделяли центрифугированием (5000 об/мин) на пищевые частицы (ПЧ), плотную эндогенную (ПЭФ), инфузорную (ИНФ) и растворимую (РФ) фракции по разработанной на кафедре физиологии и биохимии животных РГАУ-МСХА им. К.А. Тимирязева методике, основанной на изменении вязкости рубцового содержимого и скорости осаждения частиц различной плотности. В РЖ и полученных фракциях ЦРС определяли количество общего и небелкового азота — по Кьельдалю, белкового — как разницу этих показателей; в РЖ также измеряли содержание аммиачного азота микродиффузным методом в чашках Конвея и мочевины — с использованием диацетилмонооксимового реактива из набора Био-Ла-Тест («PLIVA-Lachema Diagnostika», Чехия).
Данные обрабатывали методами биометрического анализа с помощью программ Microsoft Excel и Microsoft Word. Различия между показателями считали достоверными при Р < 0,05 (7).
Результаты. Ранее мы обнаружили (8), что активность метаболических процессов в рубце достигает максимума через 3 ч после кормления животных и затем постепенно снижается по мере переваривания легкодоступных углеводов и белков. Содержание общего азота в РЖ через 3 ч составляло 281,7 мг%, на 6-й и 10-й ч — снижалось соответственно на 6,1 и 15,5 % (табл.). Выявленное увеличение этого показателя у быков на 3-й ч по сравнению с 10-м происходит за счет перехода легкорастворимых белков корма в рубцовую жидкость, а также синтетических процессов в рубце, включая образование микробиального белка.
| Динамика содержания азотистых фракций в рубцовой жидкости у быков черно-пестрой породы в зависимости от времени после кормления (Х±m) | |||
Показатель |
3 ч |
6 ч |
10 ч |
Общий азот, мг% |
281,7±12,2а |
264,6±15,1аб |
238,0±11,4б |
Белковый азот, мг% |
191,1±10,4а |
168,0±5,6аб |
154,7±10,3б |
Небелковый азот, мг% |
90,6±1,9а |
96,6±4,5а |
83,3±1,2б |
Аммиачный азот, мг% |
9,66±0,28а |
8,33±0,43б |
6,86±0,70б |
Мочевина, ммоль/л |
10,41±0,05а |
9,37±0,06б |
3,32±0,09в |
П р и м е ч а н и я. Различия статистически достоверны при Р < 0,05; наличие одинаковых верхних индексов означает отсутствие достоверных различий между показателями. |
|||
Отмеченное через 6 ч после кормления снижение концентрации общего азота в рубцовой жидкости сопровождалось увеличением доли небелкового азота и одновременным уменьшением содержания мочевины и аммиачного азота. Между концентрацией общего и белкового азота в РЖ в целом наблюдалась прямая зависимость. Наибольший показатель по белковому азоту регистрировали через 3 ч после кормления, что обусловлено активизацией метаболических процессов (в первую очередь — микробиального синтеза белка). Наименьшее содержание (83,3 мг%) небелкового азота в РЖ наблюдали через 10 ч после кормления при максимуме через 3 ч (90,6 мг%), что свидетельствует о высокой интенсивности процессов протеолиза и дезаминирования в рубце.
Количество аммиачного азота в РЖ на 3-й ч было наибольшим, на 10-й ч — снижалось до минимального. Максимальную концентрацию мочевины в РЖ также отмечали через 3 ч после кормления (см. табл.), что обусловлено ее более активным поступлением в рубец при высокой активности жвачных процессов и саливации, наименьшую — через 10 ч на фоне постепенного снижения интенсивности метаболизма.
Выявленные закономерности динамики количественного соотношения азотистых веществ в рубцовой жидкости указывают на его гомеостазирование. Такое гомеостазирование, по нашему мнению, обусловлено доступностью этих соединений в разные периоды после кормления, активностью микроорганизмов, а также процессами саливации и избирательной двусторонней проницаемостью рубцовой стенки.
Активность микробиального метаболизма (в том числе азотистого обмена) определяется наличием энергетических субстратов — углеводов. Ранее мы показали (9), что в первые 2-3 ч после кормления в качестве энергетических субстратов используются простые легкодоступные сахара, через 3-6 ч — крахмал, после 5-6 ч — клетчатка.
Эти процессы тесно связаны с величиной рН рубцового содержимого, которая, с одной стороны, также регулируется гомеостатическими механизмами, с другой — определяет жизнедеятельность микроорганизмов и активность метаболических процессов в рубце. Следовательно, величина рН может служить косвенным показателем метаболической активности. Как правило, в химусе рубца реакция среды слабокислая и зависит от интенсивности и направленности метаболических процессов: при повышении интенсивности рН несколько снижается вследствие образования большого количества кислых метаболитов (летучих жирных кислот, аминокислот и др.), при уменьшении — возрастает, приближаясь к нейтральному. Так, через 3 ч после кормления животных значение рН РЖ было наименьшим — 6,20, через 6 и 10 ч равнялось соответственно 6,45 и 6,82.
Цельное рубцовое содержимое распределялось по фракциям (ПЧ, ПЭФ, РФ и ИНФ), количественное соотношение которых (по сухому веществу) изменялось в зависимости от продолжительности пребывания корма в рубце: так, через 3, 6 и 10 ч после кормления на ПЧ, РФ, ИНФ и ПЭФ приходилось соответственно 63, 52 и 43 %; 3, 3, и 5 %; 12, 14 и 23 %; 22, 31 и 29 %. То есть доля ПЧ была наибольшей, постепенно снижаясь (до 43 %), РФ — напротив, наименьшей с минимумом в период наиболее активного пищеварения (3-6 ч после кормления) и увеличением через 10 ч, что, очевидно, связано со снижением эвакуаторной активности рубца по мере его опорожнения и подавлением микробиального синтеза. Доля ПЭФ в сухом веществе ЦРС оказалась значительной и относительно постоянной.
Известно, что доступность и переваримость белка корма зависит от его растворимости. Через 3, 6 и 10 ч после кормления животных на ПЧ приходилось соответственно 49, 34 и 20 % от общего количества азота, на РФ — 3, 2 и 3 %, на ИНФ — 10, 21 и 31 %, на ПЭФ — 38, 43 и 46 %.
Как известно, с кормом животное потребляет белки, пептиды и амиды, кроме того, в рубец поступают эндогенные азотистые соединения, в том числе мочевина. В рубце под влиянием микроорганизмов происходит существенная трансформация потребленных азотистых соединений. Часть протеинов проходит через рубец, не подвергаясь существенному протеолизу, остальные метаболизируются бактериями и простейшими, в результате роста которых накапливаются белки, имеющие высокую биологическую ценность (10). При этом распад сырого протеина корма определяется физико-химическими свойствами и составом протеина, активностью рубцовой микрофлоры и скоростью эвакуации содержимого из рубца. P. van Soest (11) предложил следующую классификацию протеиновых фракции корма: небелковый, быстро распадающийся, распадающийся со средней скоростью, медленно распадающийся и нераспадающийся азот.
В нашем опыте наблюдалось заметное снижение количества общего азота в ПЧ: с 555,57 до 357,3 мг/кг сухого вещества соответственно через 3 и 6 ч после кормления (рис. 1), поскольку значительная часть протеина корма расщепляется микрофлорой рубца и в составе РФ химуса поступает в следующие отделы пищеварительного тракта. Содержание небелкового азота в ПЧ снижалось по мере переваривания корма в рубце, при этом происходило его перераспределение между отдельными фракциями рубцового содержимого (см. рис. 1).
Поступающий в составе корма небелковый азот относительно быстро растворяется. Концентрация аминокислот в РЖ в среднем не превышает 1,4 мг%, их содержание возрастает лишь в первые часы после кормления до 6,8 мг% (12). Такое незначительное количество аминокислот одни авторы связывают с их быстрым дезаминированием и декарбоксилированием, другие — с микробиальным синтезом белка (13). В наших опытах количество общего азота в РФ заметно снижалось — с 39,61 до 16,81 мг/кг сухого вещества соответственно через 3 и 6 ч после кормления, что свидетельствует о потреблении азотистых веществ (аммиака, амидов, пептидов, аминокислот) микроорганизмами, а также о всасывании стенкой рубца и выносе некоторых из этих веществ в следующие отделы пищеварительного тракта. Рост концентрации азотистых веществ в РФ через 10 ч, вероятно, был следствием экскреции слизи и мочевины, распада белка корма в ПЧ, а также микробиального синтеза (см. рис. 1).
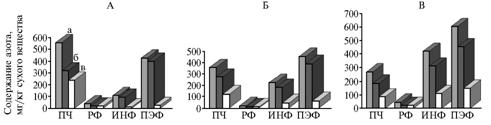 |
Рис. 1. Количество азотистых соединений во фракциях цельного рубцового содержимого у быков черно-пестрой породы через 3 (А), 6 (Б) и 10 ч (В) после кормления: ПЧ — пищевые частицы, РФ — растворимая фракция, ИНФ — инфузорная фракция, ПЭФ — плотная эндогенная фракция; а, б, в — соответственно общий, белковый и небелковый азот. |
Содержание белкового азота в РФ снижалось через 6 ч после кормления до 8,26 мг/кг сухого вещества (показатель через 3 ч — 20,08 мг/кг сухого вещества) (см. рис. 1), что объясняется либо активным поглощением растворимых белков микроорганизмами, либо поступлением эндогенного белка через 3 и 10 ч. При этом количество общего азота в ИНФ постоянно возрастало (через 3, 6 и 10 ч — соответственно 108,60; 223,21 и 419,88 мг/кг сухого вещества), что, по-видимому, связано с расширением видового состава и увеличением численности микрофлоры.
Бактерии, использующие в качестве источников азота корм, клеточный дебрис, слущивающийся рубцовый эпителий, мочевину, экскреты, фиксируются как на кормовых субстратах, так и на эпителии рубца (14, 15), после отделения которого они, по нашему мнению, могут поступать в плотную эндогенную фракцию с отмирающими эпителиальными клетками, участвуя в ее формировании наряду с муцином слюны, экскретами и другими компонентами. По нашим данным, на ПЭФ приходится 38-46 % общего азота рубцового содержимого. По данным литературы, при оценке интенсивности расщепления протеина в рубце эндогенный азот или не учитывается (в этом случае показатели занижаются), или его количество принимают равным 10-30 % от общего азота химуса (16).
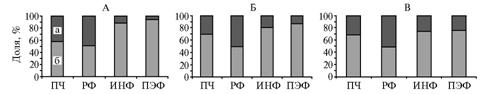 |
Рис. 2. Доля небелкового (а) и белкового (б) азота от общего количества азота во фракциях цельного рубцового содержимого у быков черно-пестрой породы через 3 (А), 6 (Б) и 10 ч (В) после кормления: ПЧ — пищевые частицы, РФ — растворимая фракция, ИНФ — инфузорная фракция, ПЭФ — плотная эндогенная фракция. |
Мы представили долю белкового и небелкового азота для каждой фракции рубцового содержимого, приняв количество общего азота в ней за 100 %. По нашим данным, в зависимости от времени после кормления в ПЭФ и ИНФ соответственно 65-75 и 70-88 % общего азота приходилось на азот белка (рис. 2). В РФ количество белкового и небелкового азота было примерно равным и не изменялось во все периоды наблюдения (см. рис. 2). Доля небелкового азота в ПЭФ возрастала с увеличением периода времени после кормления: возможно, ПЭФ, с одной стороны, связывает небелковый азот (свободные аминокислоты, короткие пептиды, амины и др.), с другой — происходит распад белковой составляющей этой фракции с высвобождением небелковых соединений.
Таким образом, в рубце жвачных животных существует сложная и весьма мощная система гомеостазирования, определяющая в течение всего периода пребывания корма в рубце функциональную стабильность последнего как полуоткрытой системы и относительно постоянное соотношение между содержанием различных азотистых соединений в рубцовой жидкости — активной фракции цельного рубцового содержимого (ЦРС). Относительная стабильность соотношения азотистых веществ достигается за счет их перемещения из одной фракции ЦРС в другую согласно закону устойчивого неравновесия, изменения активности процессов расщепления белков корма, трансформации промежуточных азотистых соединений, их всасывания и экскреции и, наконец, синтеза микробиального белка. Сложные превращения азотистых соединений в рубце энергетически и пластически обеспечивается тесно связанными с ними процессами углеводного обмена.
Л И Т Е Р А Т У Р А
1. Г е о р г и е в с к и й В.И., П о л я к о в а Е.П. Кишечный химус и процессы всасывания: новые аспекты. Тез. 2-й Междунар. конф. «Актуальные проблемы биологии в животноводстве». М., 1995: 24.2. Ш е в е л е в Н.С., Г е о р г и е в с к и й В.И., П о л я к о в а Е.П. и др. Роль эндогенных структур химуса в поддержании кишечного гомеостаза и усвоении питательных веществ. В сб.: Проблемы и перспективы развития теории питания жвачных животных на основе субстратной обеспеченности метаболизма. Боровск, 1999: 76-80.
3. Ш е в е л е в Н.С., П о л я к о в а Е.П., Л а б у н с к а я Н.А. Слизистые образования пищеварительного тракта и их роль в обмене минеральных элементов у валухов. Докл. ТСХА, 2002, 274: 353-355.
4. П о л я к о в а Е.П., К с е н о ф о н т о в Д.А, К с е н о ф о н т о в а А.А. Динамика содержания минеральных элементов во фракциях химуса разных отделов ЖКТ жвачных. Докл. ТСХА, 2000, 272: 205-210.
5. Ш е в е л е в Н.С., П о л я к о в а Е.П., К с е н о ф о н т о в Д.А. Исследование функции эндогенных слизистых образований в формировании химуса и обмене питательных веществ у животных. Докл. ТСХА/РГАУ-МСХА им. К.А. Тимирязева, 2006, 278: 714-716.
6. К а л а ш н и к о в А.П., Ф и с и н и н В.И., Щ е г л о в В.В. и др. Нормы и рационы кормления сельскохозяйственных животных. Справ. пос. М., 2003.
7. П л о х и н с к и й Н.А. Руководство по биометрии для зоотехников. М., 1970.
8. Ш е в е л е в Н.С., К а с а т к и н А.Г. Динамика процессов углеводного обмена в рубце бычков-кастратов при различном сахаро-протеиновом отношении и изменении концентрации магния в рационе. Докл. ТСХА/РГАУ-МСХА им. К.А. Тимирязева, 2000; 272: 182-186.
9. Ш е в е л е в Н.С., К а с а т к и н А.Г. Влияние кормовых и временных факторов на ферментацию углеводов в рубце быков-кастратов. Кормление с.-х. животных и кормопроизводство, 2006, 9: 29-30.
10. А л и е в А.А. Обмен веществ у жвачных животных. М., 1997.
11. V a n S o e s t P.J., S n i f f e n C.J., M a r t e n s D. R. e.a. A net protein system for cattle: The rumen submodels for nitrogen. In: Proc. Intern. symp. on protein requirements for cattle. Oklahoma, 1982: 265-279.
12. Т а р а к а н о в Б.В., Н и к о л и ч е в а Т.А. Биосинтез и метаболизм аминокислот в пищеварительном тракте лактирующих коров при разных соотношениях углеводов в рационах. Сб. науч. тр. ВНИИ физиологии, биохимии и питания с.-х. животных, 1983, 26: 80-88.
13. V a n d e r W a l t I.G., M e y e r J.H.F. Protein digestion in ruminants. S. Afr. J. Аnim. Sci., 1988, 18(1): 30-41.
14. А л и е в А.А. Достижения физиологии пищеварения сельскохозяйственных животных в ХХ веке. С.-х. биол., 2007, 2: 12-23.
15. W a l l a c e R.J., C h e n g K.J., D i n s d a l e D. e.a. An independent microbial flora of the epithelium and its role in the ecomicrobiology of the rumen. Nature, 1979, 5712(279): 424-426.
16. S m i t h R. Nitrogen metabolism in the rumen and composition and nutritive value of nitrogen compounds entering the duodenum. In: Digestion and metabolism in the ruminant. Australia, The University of New England Publishing Unit., 1975: 399-415.
METABOLISM OF FODDER NITROGEN COMPOUNDS IN RUMEN OF BULLS DEPENDING ON TIME AFTER FEEDING
N.S. Shevelev, N.A. Polozkova
In Black-and-White fistulated bulls concentration of nitrogen compounds was estimated in total rumen content, its different fractions and rumen liquid in 3, 6 and 10 hours after feeding. Comparatively constant proportion of nitrogen compounds was shown to be supported during the experiment due to active homeostatic processes.
Key words: cattle, fistulated animals, ruminants, rumen, rumen content fractions, nitrogen compounds, hoemeostasis.
ФГОУ ВПО Российский государственный |
Поступила в редакцию |
Оформление электронного оттиска
![]()
![]()




