doi: 10.15389/agrobiology.2021.1.78rus
УДК 633.11:575.21
Работа выполнена при финансовой поддержке гранта РНФ № 16-16-00011. Размножение селекционных линий до поколения F6-7 проводилось рамках госзадания ФИЦ ИЦиГ СО РАН № 0259-2021-0012, полевые испытания линий в Республике Татарстан проведены в соответствии с госзаданием ФИЦ КазНЦ РАН АААА-А18-118031390148-1.
ФЕНОТИПИЧЕСКАЯ ИЗМЕНЧИВОСТЬ СЕЛЕКЦИОННЫХ ЛИНИЙ МЯГКОЙ ПШЕНИЦЫ (Triticum aestivum L.) ПО ЭЛЕМЕНТАМ СТРУКТУРЫ УРОЖАЯ В ЭКОЛОГИЧЕСКИХ УСЛОВИЯХ ЗАПАДНОЙ СИБИРИ И ТАТАРСТАНА
А.И. СТАСЮК1 ✉, И.Н. ЛЕОНОВА1, М.Л. ПОНОМАРЕВА2, Н.З. ВАСИЛОВА2, В.П. ШАМАНИН3, Е.А. САЛИНА4
Мягкая яровая пшеница — одна из широко возделываемых зерновых культур в Российской Федерации. Селекция яровой пшеницы направлена на создание сортов, характеризующихся высокой урожайностью и качеством зерна. Основное внимание при создании новых сортов обращают на устойчивость к климатическим факторам, которые отрицательно влияют на стабильность урожайности. Что касается скрещиваний озимых и яровых сортов пшеницы, то в настоящее время нет достаточной информации о проявлении и наследовании признаков продуктивности в более старших поколениях и в различных агроэкологических условиях. В представляемой работе мы изучили изменчивость элементов структуры урожая у 55 селекционных линий мягкой пшеницы (Triticum aestivum L.) поколений F6-7, полученных от скрещивания озимых сортов мягкой пшеницы с яровыми донорами генов устойчивости к бурой ржавчине, с целью выделения генотипов, перспективных в качестве источников ценных агрономических признаков. Полевые исследования проводили в 2018 году в трех регионах — в Новосибирской и Омской областях и в Республике Татарстан. Оценивали следующие элементы структуры урожая: число продуктивных стеблей, число зерен с колоса, масса зерна с колоса, масса 1000 зерен и масса зерна с растения. Экологическую пластичность линий характеризовали с помощью показателей интенсивности и устойчивости индекса стабильности. Дисперсионный анализ, выполненный по результатам экологических испытаний, свидетельствует о достоверном (p < 0,001) влиянии генотипа, внешней среды и их взаимодействия на фенотипическое проявление всех изученных признаков. Наиболее высокий вклад генотипа показан для признаков число зерен с колоса (42,8 %) и масса 1000 зерен (57,0 %). Высокий вклад условий внешней среды установлен для продуктивной кустистости (41,8 %) и массы зерна с колоса (40,3 %). Для всех признаков существенное влияние оказывало взаимодействие генотип ½ среда, вклад которого варьировал от 25,9 % (масса 1000 зерен) до 41,0 % (масса зерна с колоса), что свидетельствует о значительной реакции генотипов на изменяющиеся условия. При полевых испытаниях наблюдалась высокая изменчивость всех признаков во всех трех климатических зонах, однако степень вариаций была различной. Усредненные показатели изученных признаков были ниже в полевых условиях Омской зоны по сравнению с Новосибирской областью и Татарстаном. Более высокие флуктуации отмечены для числа зерен с колоса (от 13,0 до 69,0), массы зерна с колоса (от 0,35 г до 2,65 г) и массы зерна с растения (от 0,15 г до 6,95 г) в условиях Татарстана по сравнению с другими регионами. Расчет индексов стабильности, устойчивости индексов стабильности и индексов интенсивности установил, что 16 генотипов из 55 входят в группу сортов интенсивного типа, 35 — полуинтенсивного и 4 — экстенсивного. Анализ методом главных координат разделил образцы на четыре основных кластера по типу стабильности и интенсивности. Молекулярный анализ селекционных линий на наличие генов устойчивости к бурой ржавчине Lr6Ai#2, LrAsp5 и LrTt2, интродуцированных соответственно от Thinopyrum intermedium (Host) Barkworth & D.R. Dewey, Aegilops speltoides Tausch и Triticum timopheevii Zhuk., показал, что 10 линий из 55 не содержат чужеродного генетического материала. Результаты маркерного анализа на присутствие генов устойчивости не коррелировали с кластеризацией образцов по типу интенсивности и стабильности. Этот факт позволяет предположить, что наличие фрагментов чужеродного генетического материала не влияет на стабильность селекционных линий. На основании проведенных исследований были отобраны генотипы, представляющие интерес в качестве источников продуктивности.
Ключевые слова: Triticum aestivum, мягкая пшеница, экологическая пластичность, хозяйственно важные признаки, продуктивность, бурая ржавчина.
Мягкая пшеница (Triticum aestivum L.) имеет большое значение во всем мире как источник продовольствия для населения и кормового зерна для сельскохозяйственных животных. Российская Федерация в настоящее время занимает лидирующие позиции по производству зерна озимой и яровой пшеницы (1). За последнее десятилетие в стране произошло увеличение посевных площадей, занятых этой культурой, увеличился валовой сбор зерна и повысилась урожайность (2, 3).
Основные производители яровой мягкой пшеницы в России — районы Поволжья, Уральский и Западно-Сибирский регионы, при этом до 40 % посевных площадей культуры приходится на Сибирский регион. Однако, несмотря на тенденцию повышения урожайности яровой пшеницы, отмечается значительное ее варьирование по годам. По данным Л.П. Силаевой и Е.В. Бариновой (3), урожайность яровой пшеницы в 2007-2017 годах в целом по стране изменялась от 9,5 до 18,5 ц/га. Существенное колебание урожая показано как для стародавних, так и для современных яровых сортов пшеницы, выращиваемых в Западной Сибири (4-6).
Генетическая основа сорта — базовый фактор, определяющий урожайность. В последние годы с появлением насыщенных молекулярных карт и развитием технологий высокопроизводительного гено- и фенотипирования достигнут существенный прогресс в изучении генетической архитектуры признаков урожая и его компонентов у мягкой пшеницы. Идентифицированы главные и минорные локусы, которые вносят вклад в фенотипическое проявление продуктивной кустистости, числа зерен и массы зерна с колоса, массы зерна с растения, параметров корневой системы и др. (7-9). Определены локусы количественных признаков (QTLs) и гены-кандидаты, детерминирующие урожайность в условиях абиотических стрессовых факторов (10-13). Однако реализация генетического потенциала пшеницы в значительной степени зависит от факторов внешней среды. Поэтому на определенных этапах получения новых форм необходимо изучение их фенотипической пластичности и адаптивности в различных почвенно-климатических условиях.
Озимая пшеница отличается от яровой более высокой продуктивной кустистостью, массой зерна с колоса и массой 1000 зерен, которые вносят основной вклад в урожайность сортов. Однако яровая пшеница в отличие от озимой характеризуется высокими показателями качества зерна и клейковины и более засухоустойчива (14-16). C середины XX века для получения новых яровых и озимых сортов пшеницы с высокой урожайностью и устойчивостью к болезням используется стратегия объединения генетических пулов этих культур. Несмотря на то, что у большинства сортов озимой пшеницы комбинационная способность при скрещивании с яровой пшеницей характеризуется как средняя либо слабая, показано, что в ряде случаев гибридизация может дать желательные генотипы (17, 18). Общую комбинационную способность (ОКС) популяций от скрещивания озимых и яровых образцов изучали в основном на стадии F1 для выявления эффекта гетерозиса и отбора желательных генотипов с более высокими показателями по хозяйственно важным признакам (19, 20). Оценку ОКС в поколениях F1-F2 проводили в основном на гибридных популяциях, полученных на основе гибридизации либо озимых, либо яровых сортов. Авторы этих исследований утверждают, что поколение F1 может использоваться для начальной стадии оценки ОКС и отбора доноров. Так, изучение комбинационной способности по признаку «зимостойкость» у 30 диаллельных гибридов поколения F1, полученных от скрещивания 6 озимых сортов пшеницы, позволило выявить генотипы, характеризующиеся высокими эффектами ОКС и участием доминантных аллелей в увеличении признака (21). На основании полученных результатов был сделан вывод, что для отбора уникальных генотипов с усиленной зимостойкостью необходима проверка гибридов поколений F4-F6. Исследования для признаков продуктивности, выполненное с использованием F1-F2 гибридов яровых и озимых сортов, указывают на значительный вклад генов с аддитивным типом действия, что, по мнению авторов, позволяет начинать отбор доноров в ранних поколениях (22, 23). Что касается скрещиваний озимых и яровых сортов пшеницы, то в настоящее время нет достаточной информации о проявлении и наследовании признаков продуктивности в более старших поколениях и в различных агроэкологических условиях.
Ранее нами для расширения генетического разнообразия мягкой пшеницы по генам устойчивости к болезням были созданы селекционные линии на основе гибридизации озимых сортов и яровых доноров генов резистентности (24, 25).
В настоящем исследовании выполнена оценка этих селекционных линий по элементам структуры урожая в разных экологических условиях.
Целью работы было изучение экологической пластичности образцов продвинутых поколений и выделение перспективных генотипов в качестве источников признаков продуктивности для селекции.
Методика. Исследования проводили на 55 линиях яровой мягкой пшеницы поколений F6-7, полученных от скрещивания озимых сортов Новосибирская 3 (Н3), Новосибирская 40 (Н40), Филатовка и Бийская озимая с яровыми донорами генов устойчивости к бурой ржавчине — сортом Тулайковская 10 (Т10) и линиями 5366-180 и 21-4, полученными соответственно на сортах Саратовская 29 и Новосибирская 29. Сорт Т10 содержит ген Lr6Ai#2 от Thinopyrum intermedium (Host) Barkworth & D.R. Dewey, линия 8366-180 — ген LrTt2 от Triticum timopheevii Zhuk., линия 21-4 — ген LrAsp5 от Aegilops speltoides Tausch.(24, 26). Линии были отобраны по срокам колошения и параметрам продуктивности в поколениях F3-5 и размножены до поколений F6-7.
ДНК выделяли из листьев растений поколения F2. Фрагмент листа (2-3 см) помещали в пробирку объемом 2 мл с матриксами для измельчения растительных тканей Lysing Matrix Z («MP Biomedicals», США), добавляли 700 мкл буфера для лизиса (1 мл 1 М Tris-HCl, 1 мл 5 М NaCl, 1 мл 0,5 М EDTA, 0,625 мл 20 % SDS и 0,04 г Na2S2O5), измельчали на гомогенизаторе FastPrep-24 («MP Biomedicals», США) и инкубировали на водяной бане при 60 °С в течение 30 мин. Затем добавляли 700 мкл смеси хлороформ:изоамиловый спирт (24:1) и центрифугировали 25 мин при 12000 об/мин. Верхнюю фракцию помещали в пробирку объемом 2 мл, добавляли 1,4 мл 96 % охлажденного этанола (-20 °С) и центрифугировали 15 мин при 4500 об/мин. К осадку добавляли 700 мкл 70 % этанола и центрифугировали 5 мин при 4500 об/мин. Супернатант удаляли, осадок высушивали на воздухе и растворяли в 50 мкл буфера ТЕ.
Для определения генотипов, содержащих ген Lr6Ai#2 от Th. intermedium, использовались разработанные нами праймеры МF2 (5´-GATGTCG-AGGAGCATTTTC-3´), MR1r2 (5´-GTGGTAGATTACTAGAGTTCAAGTG-3´) и MR4 (5´-CGAATAGTTATACTAGGAGTAC-3´) (Патент № RU 2598275 C1, 2015). Эти праймеры доминантные, и для выявления гомозиготных и гетерозиготных по этому гену растений необходимо использовать две пары — МF2/MR1r2 и MF2/MR4. Для идентификации интрогрессированных фрагментов от T. timopheevii, содержащих генLrTt2, использовали микросателлитный маркер Xbarc232 (F — 5´-CGCATCCAACCATCCCCACCCAACA-3´, R — 5´-CGCAGTAGATCCACCACCCCGCCAGA-3´) (27). Транслокации от Ae. speltoides, содержащие ген LrAsp5, определяли с помощью пары праймеров P1/P5 (P1 — 5´-TACCCCTGCTACCAGTGCGC-3´, P5 — 5´-GGCCA-ACCCTACACCCCAAG-3´) (Патент № RU 2535985 C1, 2013).
ПЦР проводили на амплификаторе T-100 («Bio-Rad», США) в общем объеме реакционной смеси 20 мкл, содержащей ДНК (50-100 нг), 10× буфер для Taq полимеразы (650 мМ Tris-HCl, pH 8,9; 160 мМ (NH4)2SO4, 25 мМ MgCl2, 0,01 % Tween 20), по 0,25 мМ каждого dNTP, по 1 нг каждого праймера, 1 мкл Taq полимеразы (1 ед. активности/мкл), H2O (до конечного объема). Условия полимеразной цепной реакции (ПЦР) для выявления генов описаны ранее (24). Продукты ПЦР разделяли с помощью электрофореза в 1,5 % агарозном геле, содержащем бромистый этидий.
Опытные испытания образцов проводили в полевых условиях Новосибирской и Омской областей и в Республике Татарстан (г. Казань) в 2018 году. Почва опытного участка в Новосибирской области — чернозем выщелоченный, в Омской — лугово-черноземная малогумусовая среднесуглинистая, в Татарстане — хорошо окультуренная серая лесная. Полевой опыт закладывали в 2 повторностях систематическим методом на делянках шириной 1 м, по 60 зерен в ряду.
Урожай собирали в снопы, после чего оценивали число продуктивных стеблей, число зерен в колосе, массу зерна с колоса и с растения, массу 1000 зерен. Структурный анализ выполняли для 20 растений каждого образца.
Экологическую пластичность линий оценивали на основании признака «масса зерна с растения», используя показатели индекса интенсивности (И), индекса стабильности (ИС) и устойчивости индекса стабильности (У) согласно методическим указаниям (28). Величину И, показывающую реакцию линий на фон, определяли, как отношение разности массы зерна с растения по двум крайним ее значениям для каждой линии к средней массе для всех линий на всех фонах:

Оптимальным фоном считали тот регион, где показатель массы зерна с растения у конкретного образца был наибольшим, лимитированным — регион, в котором масса зерна с растения была наименьшей. Кроме того, для отнесения линии к конкретному типу интенсивности был рассчитан обобщенный показатель интенсивности для гипотетической линии (И) по средним характеристикам всех линий на испытываемых фонах и наименьшая существенная разность (НСР) частных средних по остаточной дисперсии после проведения двухфакторного дисперсионного анализа (29). Классификацию линий по степени отзывчивости на агрофон проводили по соотношениям: Илинии > И + НСР — интенсивная, И - НСР ≤ Илинии ≤ И + НСР — полуинтенсивная, Илинии < И - НСР — экстенсивная. Индекс стабильности (ИС), характеризующий проявление гомеостатических реакций линий в разных условиях среды, оценивали, как отношение квадрата средней величины массы зерна с растения у образца на конкретном фоне к величине среднеквадратичного отклонения этого показателя в определенных условиях:
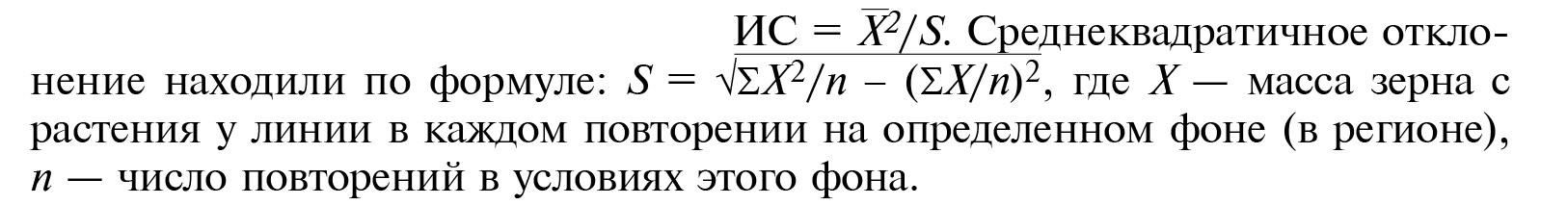
Показатель устойчивости индекса стабильности (У), характеризующий адаптивную способность линии, определяли по формуле:

Статистическую обработку результатов проводили с помощью программных пакетов MS Excel 2016 и STATISTICA v. 10.0 («StatSoft, Inc.», США). Оценивали среднее (M), минимальные (min) и максимальные (max) значения признаков, медиану (Me) и стандартную ошибку средней (±SEM). Вклад факторов рассчитывали на основании среднего квадрата отклонений (MS). За достоверный принимали уровень значимости p < 0,05. Анализ методом главных координат выполняли с использованием программы PAST v. 3.15 (30).
Результаты. Западносибирский регион относится к территориям с рискованным земледелием и включает различные климатические зоны, о чем свидетельствуют метеорологические данные по Новосибирской и Омской областям в 2018 году. Погодные условия в Новосибирской области в период вегетации отличались от среднемноголетних, средняя температура в мае была значительно ниже (6,9 °С), май и июнь характеризовались обильными осадками, превышающими норму почти в 3 раза. В Омской области средняя температура не отличалась от среднемноголетней, осадков за период май-сентябрь выпало 259 мм, при этом их наибольшее количество пришлось на вторую половину периода вегетации. В Татарстане в мае, наоборот, наблюдался дефицит осадков, в остальные месяцы они выпадали неравномерно; в июне среднесуточная температура воздуха была ниже нормы, в остальной период — на 2-3 °С выше нормы.
Характеристика 55 изученных линий по происхождению и структуре урожая при экологических испытаниях представлена в таблице 1 (см. http://www.agrobiology.ru).
Для оценки экологической пластичности использовали один из основных признаков продуктивности — массу зерна с растения, которая, по сути, служит показателем урожайности растения. Расчет показателей экологической пластичности позволил разделить изученные линии по интенсивности и адаптивности (табл. 2).
Факторный дисперсионный анализ, выполненный на основании результатов полевых испытаний, показал достоверное влияние (p < 0,001) генотипа, условий внешней среды и их взаимодействия на фенотипическое проявление признаков (табл. 3). Вклад фактора «генотип» варьировал в зависимости от признака, при этом наиболее высокий вклад отмечали для числа зерен с колоса (42,8 %) и массы 1000 зерен (57,0 %). Влияние генотипических факторов на проявление фенотипической изменчивости продуктивной кустистости и массы зерна с растения было менее значительным по сравнению с факторами внешней среды (соответственно 20,4 и 22,1 %). Высоким вкладом в фенотипическую изменчивость всех изученных признаков характеризовалось взаимодействие генотип × среда, что свидетельствует о значительной реакции генотипов на почвенные и погодные условия.
Существенный вклад фактора «генотип × среда» описан для поколения F1, полученного от скрещивания озимых и яровых сортов пшеницы (19). Однако авторы при этом установили, что эффект гетерозиса зависел не столько от условий выращивания гибридов, сколько от генотипа сортов, использованных для гибридизации. По данным многих исследований, колебание погодных условий в регионах испытаний, переувлажнение в начале вегетации, пониженные температуры приводят к значительным вариациям урожая и его компонентов (31, 32). Полученные нами результаты согласуются с данными литературы, которые свидетельствуют о неодинаковом вкладе генотипа и внешней среды в формирование урожая у мягкой пшеницы (6, 33, 34). Доля генотипа в фенотипической изменчивости урожая значительно меньше по сравнению с факторами внешней среды и агротехническими факторами, о чем свидетельствуют результаты большинства публикаций (35-37). В ряде работ показано, что реализация генетического потенциала сортов зависит не столько от влияния климатических факторов, сколько от агротехнологических мероприятий и использования интенсивных технологий, снижающих негативные эффекты внешней среды, что приводит к доминирующему влиянию генотипа (38-40).
Для обобщенной оценки изменчивости признаков урожайности в каждом из трех регионов мы провели статистическую обработку данных полевых испытаний суммарно для 55 линий без учета их происхождения. Как видно из рисунка 1, значительная изменчивость всех признаков наблюдалась при выращивании образцов во всех трех климатических зонах, однако степень вариаций была различной. Для всех признаков полученное усредненное значение было ниже в полевых условиях Омской области. Наиболее контрастный размах колебаний наблюдался по числу продуктивных стеблей в Новосибирской области (1-6 стеблей) по сравнению с Омской (1-2 стебля) и Татарстаном (1-4 стебля). Более существенные флуктуации отмечались для числа зерен с колоса, массы зерна с колоса и массы зерна с растения в условиях Татарстана по сравнению с другими регионами (см. рис. 1).
В настоящее время для расчета параметров экологической пластичности используются различные методики, предложенные S.A. Eberhart и W.A. Russell, G.C.C. Tai, Р.А. Удачиным и А.П. Головоченко и др. (28, 41, 42). Наиболее универсальной признана методика S.A. Eberhart и W.A. Russell (41), которая применяется большинством исследователей для оценки экологической пластичности и адаптивности возделываемых сортов и на конечных этапах селекционных испытаний. Однако указанный способ расчетов не позволяет получить адекватные и статистически достоверные результаты в тех случаях, когда экспериментальные образцы находятся на ранних стадиях формообразовательного процесса либо число полевых испытаний ограничено. Для оценки экологической пластичности новых селекционных линий мы использовали методику Р.А. Удачина и А.П. Головоченко (28), с помощью которой можно определять тенденцию формирования пластичности на начальных стадиях создания образцов. Эффективность метода для получения предварительной информации о пластичности сортов в условиях ограниченного числа агрофонов и сезонов испытаний была подтверждена в ряде работ (6, 43, 44).
По результатам испытаний 55 линий для каждой из трех климатических зон были рассчитаны индексы стабильности (ИС), а также индексы интенсивности (И) и показатели устойчивости индексов стабильности (У) (см. табл. 2). Параметры интенсивности позволяют определить степень отзывчивости генотипов на изменение условий возделывания растений. Индекс стабильности показывает, насколько стабильно линия способна реализовать свой потенциал в разных условиях среды. Линии с большими индексами более стабильны, то есть лучше приспособлены к изменяющимся условиям. Исследование установило, что 16 генотипов входят в группу сортов интенсивного типа, 35 — полуинтенсивного и 4 — экстенсивного.
Анализ методом главных координат, выполненный на основании рассчитанных параметров экологической пластичности, дополнительно разделил изученные селекционные линии на четыре основных кластера (рис. 2). В первый кластер вошли 9 линий, для 8 из которых характерен низкий коэффициент устойчивости (У), свидетельствующий о нестабильном типе линий. Исключением была линия № 50, которая относится к стабильному типу. Среди генотипов этой группы присутствуют образцы как интенсивного, так и полуинтенсивного типа. Кластеры 2 и 3 включают соответственно 13 и 11 образцов стабильного типа, при этом все линии, за исключением образца № 53, относятся к полуинтенсивным. Отличительная особенность кластера 3 заключается в более высоких показателях устойчивости индекса стабильности по сравнению с образцами кластера 2. И, наконец, кластер 4 объединяет 15 стабильных линий преимущественно интенсивного типа. Анализ методом главных координат также показал, что кластеры формируются из генотипов, происходящих из различных комбинаций скрещивания. Семь линий (№№ 8, 29, 30, 40, 41, 46, 55) не были отнесены ни к одному из кластеров, при этом линия № 8 существенно отличалась от всех образцов очень низким индексом стабильности (ИС = -138,54) и высоким коэффициентом интенсивности (И = 231,49). Несмотря на то, что комбинации Бийская озимая × Тулайковская 10 и Филатовка × 5366-180 представлены всего двумя образцами каждая, результаты оценки этих линий не влияют на группировку образцов.
Генотипирование образцов маркерами генов Lr6Ai#2, LrTt2 и LrAsp5 установило наличие фрагментов амплификации с праймерами МF2/MR1r2 и MF2/MR4 у линий №№ 11-21 (Филатовка × Тулайковская 10), позволяющее предположить, что они несут транслокацию от Th. intermedium (рис. 3). Чужеродные транслокации с генами устойчивости мы не выявили у линий № 1 и № 2 (Бийская озимая × Тулайковская 10), а также у линий №№ 5, 6, 7, 8, 9 и 10 (Филатовка × Тулайковская 10).Присутствие фрагментов генома от Ae. speltoides установлено с помощью праймеров Р1/Р5 у линий №№ 22-55, полученных с участием сортов Новосибирская 3 и Новосибирская 40. ПЦР-анализ двух участвующих в исследовании линий — № 3 и № 4 (Филатовка × линия 5366-180) с помощью микросателлитного маркера Xbarc232 показал отсутствие у этих линий интрогрессий от T. timopheevii. Результаты маркерного анализа на присутствие генов устойчивости не коррелировали с кластеризацией образцов по типу интенсивности и стабильности. Полученные результаты позволяют сделать заключение, что наличие чужеродного генетического материала в генотипической среде озимых сортов-реципиентов не оказывает влияния на фенотипическое проявление признаков урожайности и параметры экологической пластичности.
До настоящего времени не получено однозначных результатов о влиянии чужеродных транслокаций с генами устойчивости на проявление признаков продуктивности. Известно, что чужеродный генетический материал, входящий в состав протяженных интрогрессированных фрагментов, может оказывать как негативный, так и положительный эффект на такие признаки, как число зерен в колосе, масса зерна с колоса и масса 1000 зерен (45, 46). Подобные эффекты продемонстрированы на примере пшенично-ржаных транслокаций 1BL/1RS и 1AL/1RS, широко используемых в мире для создания сортов с комплексной устойчивостью к грибным болезням. Разнонаправленные эффекты в отношении компонентов урожайности при интрогрессии генетического материала различных видов пырея Thinopyrum ssp., эгилопсов Aegilops ssp. и T. timopheevii в геном мягкой пшеницы в значительной степени связаны с генотипической средой сорта-реципиента, о чем свидетельствуют данные литературы (47, 48).
Среди образцов, входящих в кластеры 2 и 3, мы отобрали 10 стабильных селекционных линий полуинтенсивного типа, выделяющихся более высокой массой зерна с растения и массой 1000 зерен (табл. 4). Эти линии будут вовлечены в следующий этап испытаний для оценки перспективности в качестве источников продуктивности.
Таким образом, в результате изучения набора селекционных линий мягкой пшеницы поколений F6-7 в трех эколого-географических регионах было установлено, что вклад генотипа и условий внешней среды в фенотипическую изменчивость зависит от изучаемого признака. Отмечено высокое влияние взаимодействия генотип × среда (25,9-41,0 %) на все компоненты продуктивности. Проведенный анализ экологической пластичности показал, что линии группируются в четыре основных кластера по типу стабильности и интенсивности, при этом на кластеризацию образцов не влияет происхождение генотипов из комбинаций скрещивания и наличие фрагментов чужеродного материала от Thinopyrum intermedium и Aegilops speltoides. На основании оценки экологической пластичности отобраны стабильные генотипы полуинтенсивного типа (№№ 14, 15, 19, 20, 24, 26, 32, 42, 47 и 49) для дальнейших испытаний при различных технологиях возделывания в условиях одного региона.
2. Параметры экологической пластичности 55 изученных образцов пшеницы F6-7 от разных комбинаций |
||||||
№ линии |
И, % |
У, % |
ИС |
Тип |
||
Новосибирск |
Омск |
Казань |
||||
Озимый сорт Бийская озимая × яровой сорт Тулайковская 10 |
||||||
1 |
130,27 |
-3,3 |
7,35 |
2,62 |
8,55 |
Ин/С |
2 |
92,95 |
48,1 |
7,72 |
4,74 |
5,71 |
Ин/С |
Озимый сорт Филатовка × яровая линия 5366-180 |
||||||
3 |
108,84 |
-2,5 |
5,09 |
2,32 |
8,20 |
Ин/С |
4 |
71,00 |
51,3 |
5,59 |
6,48 |
8,38 |
П-Ин/С |
Озимый сорт Филатовка × яровой сорт Тулайковская 10 |
||||||
5 |
99,52 |
-52,7 |
14,15 |
7,02 |
15,78 |
Ин/НС |
6 |
22,78 |
84,4 |
6,36 |
5,46 |
5,52 |
П-Ин/С |
7 |
89,56 |
54,6 |
7,73 |
9,31 |
10,34 |
Ин/С |
8 |
231,49 |
-138,5 |
5,78 |
2,66 |
16,35 |
Ин/НС |
9 |
109,09 |
4,1 |
6,93 |
3,88 |
9,38 |
Ин/С |
10 |
22,95 |
83,2 |
5,81 |
4,84 |
5,46 |
П-Ин/С |
11 |
75,54 |
-24,2 |
9,36 |
2,23 |
4,32 |
П-Ин/НС |
12 |
97,12 |
31,1 |
8,21 |
4,30 |
4,26 |
Ин/С |
13 |
91,31 |
6,5 |
7,73 |
2,36 |
4,54 |
Ин/С |
14 |
41,58 |
30,5 |
4,03 |
4,71 |
8,02 |
П-Ин/С |
15 |
44,98 |
72,3 |
6,86 |
6,10 |
5,27 |
П-Ин/С |
16 |
60,83 |
11,7 |
7,05 |
1,98 |
4,93 |
П-Ин/С |
17 |
84,00 |
-36,4 |
7,72 |
3,17 |
11,00 |
Ин/НС |
18 |
59,79 |
21,6 |
5,00 |
2,31 |
6,81 |
П-Ин/С |
19 |
30,71 |
74,0 |
5,26 |
5,82 |
4,32 |
П-Ин/С |
20 |
39,65 |
64,8 |
3,47 |
2,77 |
4,79 |
П-Ин/С |
21 |
117,18 |
13,6 |
6,65 |
3,03 |
7,99 |
Ин/С |
Озимый сорт Новосибирская 3 × яровая линия 21-4 |
||||||
22 |
33,05 |
52,6 |
4,68 |
2,64 |
5,36 |
П-Ин/С |
23 |
72,26 |
30,9 |
5,58 |
3,76 |
7,73 |
П-Ин/С |
24 |
34,73 |
75,0 |
4,10 |
3,95 |
5,38 |
П-Ин/С |
25 |
31,93 |
85,4 |
5,14 |
4,77 |
4,30 |
П-Ин/С |
Озимый сорт Новосибирская 40 × яровая линия 21-4 |
||||||
26 |
42,53 |
67,5 |
6,34 |
7,09 |
5,22 |
П-Ин/С |
27 |
19,85 |
44,3 |
6,64 |
3,44 |
6,04 |
П-Ин/С |
28 |
22,78 |
53,6 |
6,17 |
7,60 |
4,94 |
П-Ин/С |
29 |
50,62 |
91,0 |
5,81 |
5,62 |
6,14 |
П-Ин/С |
30 |
13,53 |
68,4 |
4,36 |
2,55 |
4,20 |
Э/С |
31 |
36,58 |
66,8 |
4,10 |
2,23 |
4,14 |
П-Ин/С |
32 |
55,58 |
44,5 |
4,40 |
3,32 |
6,51 |
П-Ин/С |
33 |
66,84 |
46,0 |
6,21 |
3,76 |
6,87 |
П-Ин/С |
34 |
63,17 |
-27,9 |
10,36 |
3,54 |
3,01 |
П-Ин/НС |
35 |
54,90 |
39,1 |
7,20 |
3,70 |
4,58 |
П-Ин/С |
36 |
82,37 |
48,8 |
7,99 |
6,19 |
9,13 |
Ин/С |
37 |
28,07 |
85,9 |
3,45 |
4,18 |
3,37 |
П-Ин/С |
38 |
52,26 |
17,6 |
9,28 |
4,55 |
4,72 |
П-Ин/С |
39 |
38,26 |
54,2 |
5,16 |
2,53 |
4,30 |
П-Ин/С |
40 |
16,54 |
96,4 |
4,60 |
4,40 |
4,42 |
Э/С |
41 |
18,26 |
5,1 |
9,03 |
3,58 |
3,90 |
Э/С |
42 |
36,76 |
76,2 |
5,55 |
4,19 |
4,43 |
П-Ин/С |
43 |
42,93 |
-18,2 |
11,19 |
4,41 |
5,88 |
П-Ин/НС |
44 |
88,26 |
-46,2 |
12,28 |
3,89 |
7,14 |
Ин/НС |
45 |
81,10 |
23,5 |
7,45 |
3,06 |
5,54 |
Ин/С |
46 |
42,57 |
85,0 |
4,39 |
4,05 |
4,91 |
П-Ин/С |
47 |
46,41 |
53,6 |
6,97 |
4,31 |
5,97 |
П-Ин/С |
48 |
92,70 |
28,8 |
8,24 |
5,54 |
4,15 |
Ин/С |
49 |
33,38 |
32,7 |
9,54 |
5,87 |
5,68 |
П-Ин/С |
50 |
66,14 |
-11,6 |
10,26 |
3,86 |
3,90 |
П-Ин/С |
51 |
54,85 |
-16,0 |
11,38 |
4,84 |
4,72 |
П-Ин/НС |
52 |
96,35 |
37,2 |
4,49 |
5,19 |
8,09 |
Ин/С |
53 |
15,06 |
57,7 |
5,31 |
2,88 |
4,72 |
Э/С |
54 |
64,79 |
-15,8 |
9,56 |
3,90 |
2,91 |
П-Ин/НС |
55 |
45,89 |
89,2 |
5,77 |
5,15 |
5,55 |
П-Ин/С |
Примечение. И — индекс интенсивности, У — устойчивость индекса стабильности, ИС — индекс стабильности; |
||||||
3. Факторный дисперсионный анализ структуры урожая у 55 изученных образцов пшеницы F6-7 от разных комбинаций скрещивания озимых форм с яровыми при испытаниях в Новосибирской, Омской областях и Республике Татарстан (для каждого образца n = 20, 2-кратная биологическая повторность, 2018 год) |
||||
Источник вариации Признак, параметр |
Генотип |
Регион |
Генотип × регион |
Ошибка |
Продуктивная кустистость: |
|
|
|
|
df MS |
54 4,06 |
2 225,13 |
108 3,77 |
0,86 |
F |
4,7* |
260,6* |
4,4* |
|
вклад, % |
20,4 |
41,8 |
35,9 |
|
Число зерен с колоса: |
|
|
|
|
df MS |
54 774 |
2 10451 |
108 325 |
80 |
F |
9,7* |
131,5* |
4,1* |
|
вклад, % |
42,8 |
21,4 |
35,9 |
|
Масса зерна с колоса: |
|
|
|
|
df MS |
54 1,22 |
2 23,11 |
108 0,72 |
0,16 |
F |
7,6* |
144,7* |
4,5* |
|
вклад, % |
34,6 |
24,4 |
41,0 |
|
Масса 1000 зерен: |
|
|
|
|
df MS |
54 422 |
2 3514 |
108 94 |
28 |
F |
15,3* |
127,7* |
3,4* |
|
вклад, % |
57,0 |
17,6 |
25,4 |
|
Масса зерна с растения: |
|
|
|
|
df MS |
54 9,33 |
2 458,70 |
108 7,94 |
1,43 |
F |
6,5* |
320,2* |
5,5* |
|
вклад, % |
22,1 |
40,3 |
37,6 |
|
Примечение. df — число степеней свободы, MS — средний квадрат; F — критерий Фишера. Для каждого образца n = 20, 2-кратная биологическая повторность. * Вклад фактора статистически значим при p < 0,001. |
||||
4. Характеристика стабильных селекционных линий пшеницы полуинтенсивного типа от разных |
||||||||
№ линии |
Элемент структуры урожая, M±SEM |
Наличие генов устойчивости |
||||||
число, шт. |
масса, г |
|||||||
продуктивных стеблей |
зерен в колосе |
зерен с колоса |
зерен с растения |
1000 зерен |
Lr6Ai#2 |
LrTt2 |
LrAsp5 |
|
14 |
1,50±0,09 |
43,28±1,43 |
1,60±0,06 |
2,18±0,13 |
37,34±1,07 |
+ |
- |
- |
15 |
1,75±0,10 |
37,63±0,96 |
1,49±0,05 |
2,23±0,13 |
38,81±0,57 |
+ |
- |
- |
19 |
1,85±0,12 |
38,00±1,59 |
1,50±0,07 |
2,18±0,14 |
37,62±0,85 |
+ |
- |
- |
20 |
1,83±0,15 |
37,48±1,41 |
1,45±0,07 |
2,27±0,19 |
37,71±0,62 |
+ |
- |
- |
24 |
2,02±0,13 |
33,25±1,04 |
1,22±0,04 |
2,11±0,14 |
35,86±0,96 |
- |
- |
+ |
26 |
1,77±0,13 |
41,13±1,11 |
1,57±0,05 |
2,30±0,15 |
36,74±0,60 |
- |
- |
+ |
32 |
1,60±0,10 |
39,33±1,56 |
1,52±0,07 |
2,09±1,52 |
37,78±0,75 |
- |
- |
+ |
42 |
2,22±0,14 |
34,85±1,13 |
1,30±0,05 |
2,34±0,16 |
35,57±0,64 |
- |
- |
+ |
47 |
2,12±0,13 |
39,40±0,98 |
1,50±0,05 |
2,62±0,17 |
35,26±0,62 |
- |
- |
+ |
49 |
2,05±0,12 |
39,88±0,97 |
1,56±0,04 |
2,62±0,14 |
37,16±0,64 |
- |
- |
+ |
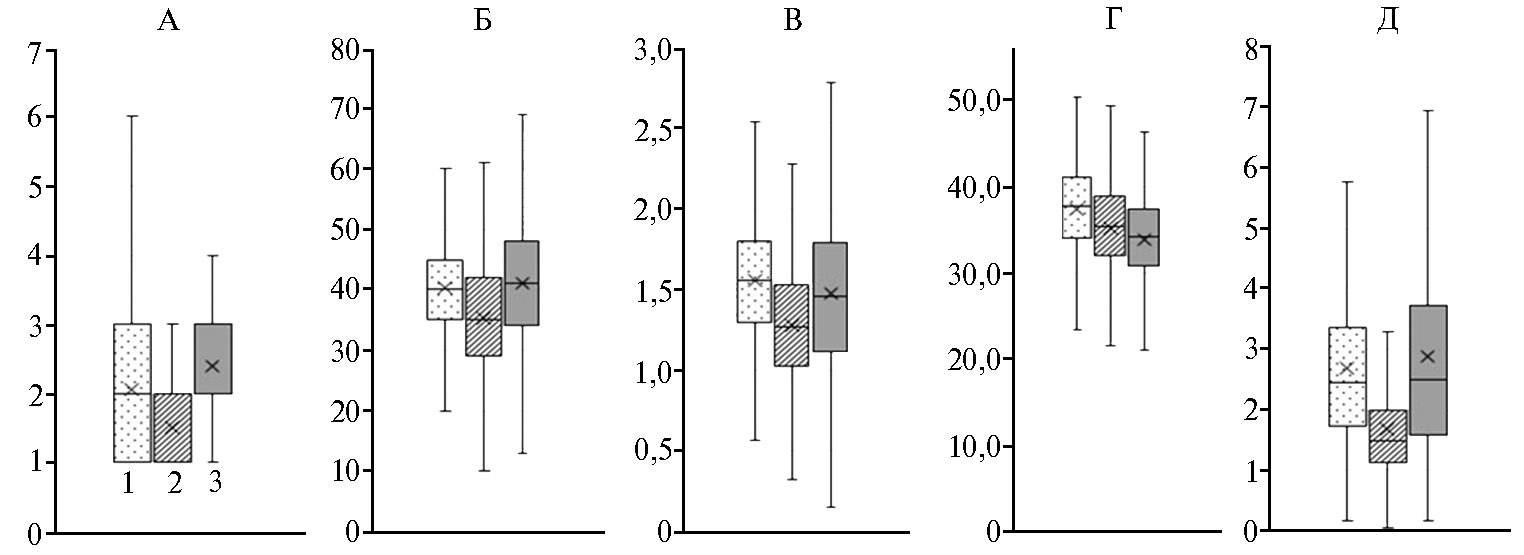
Рис. 1. Статистический анализ вариабельности элементов структуры урожая у 55 изученных образцов пшеницы F6-7 без учета их происхождения при испытаниях в Новосибирской области (1), Омской области (2) и Республике Татарстан (3) (2018 год): А — число продуктивный стеблей, шт.; Б — число зерен с колоса, шт.; В — масса зерен с колоса, г; Г — масса 1000 зерен, г; Д — масса зерен с растения. Усы обозначают минимальное (min) и максимальное (max) значения признака, горизонтальная черта в боксе — медиана (Me),знаком «×» отмечено среднее значение признака. Образцы получены в комбинациях скрещиваний озимых сортов с яровыми формами. Для каждого образца n = 20, 2-кратная биологическая повторность.
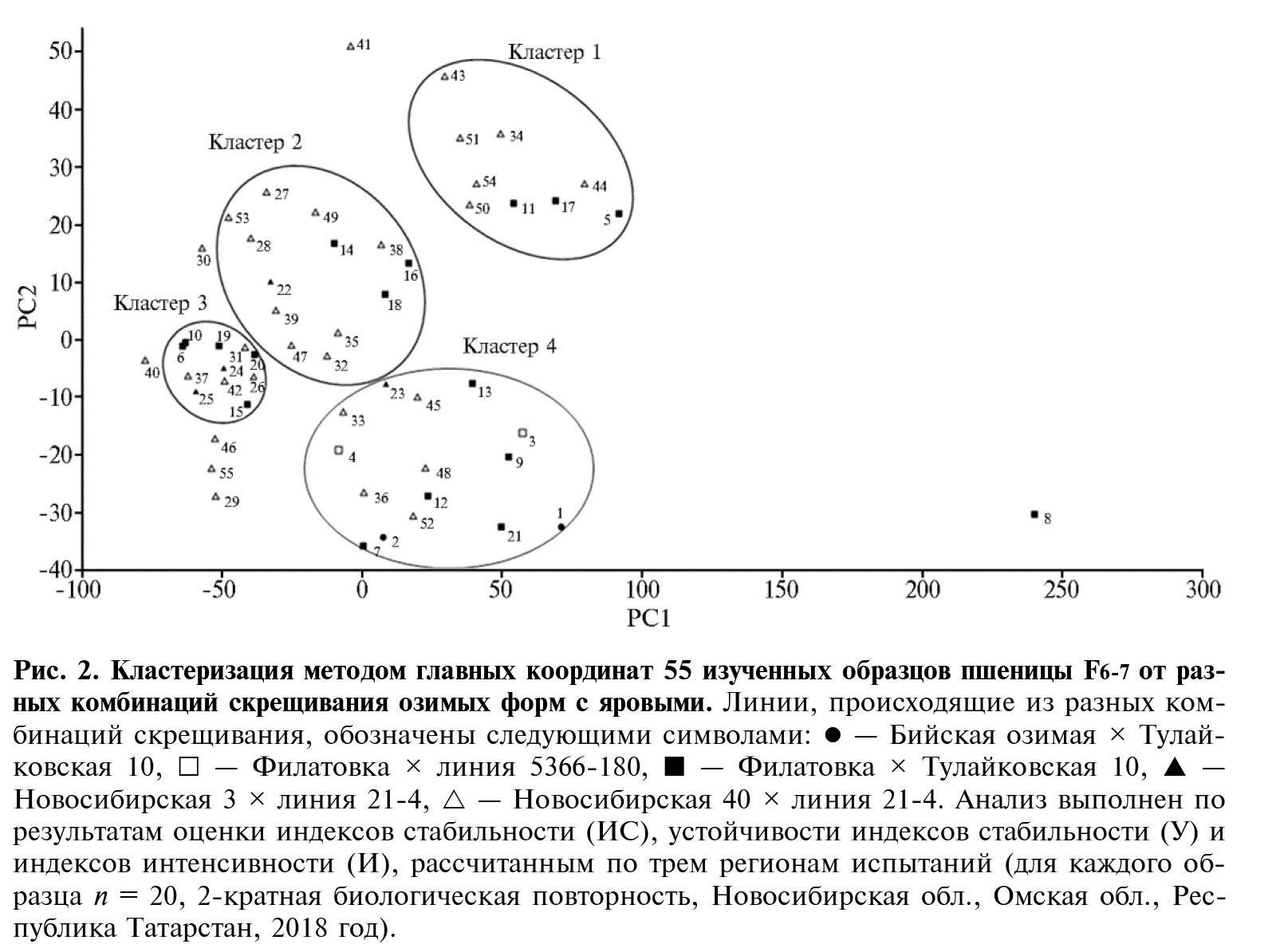
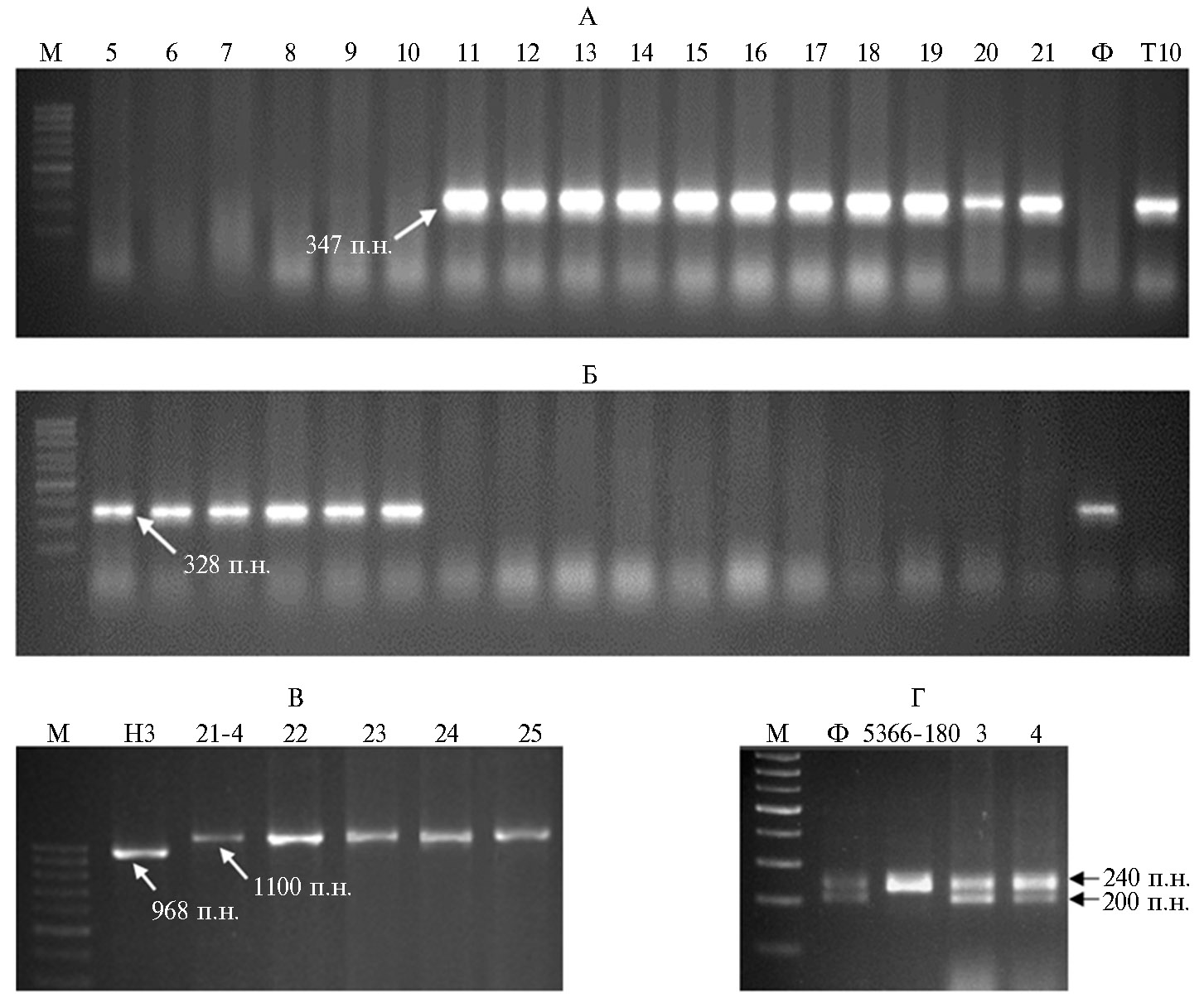
Рис. 3. Пример идентификации генотипов с генами устойчивости к бурой ржавчине среди 55 изученных образцов пшеницы F6-7 от разных комбинаций скрещивания озимых форм с яровыми. Праймеры MF2/MR1r2 (А) и MF2/MR4 (Б) выявляют ген Lr6Ai#2, праймеры P1/P5 (В) выявляют ген LrAsp5, маркер Xbarc232 (Г) выявляет ген LrTt2;5-21 — линии, полученные от скрещивания сортов Филатовка × Тулайковская 10, Ф — сорт Филатовка, Т10 — сорт Тулайковская 10; 22-25 — линии, полученные от скрещивания Новосибирская 3 × линия 21-4, Н3 — сорт Новосибирская 3, 21-4 — родительская линия 21-4; 3, 4 — линии, полученные от скрещивания Филатовка × линия 5366-180. М — маркер длины фрагментов ДНК (100 bp ladder, ЗАО «Биосан», г. Новосибирск). Стрелки указывают на диагностические фрагменты 347 п.н., показывающие наличие транслокаций от Thinopyrum intermedium, и 328 п.н., свидетельствующие об отсутствии такой транслокаций. Фрагмент 1100 п.н. показывает наличие транслокаций от Aegilops speltoides, а 968 п.н. — ее отсутствие. Фрагмент 240 п.н. указывает на наличие транслокаций от Triticum timopheevii, амплификация двух фрагментов 200 и 240 п.н. свидетельствует об отсутствии транслокаций.
ЛИТЕРАТУРА
- FAOSTAT. Режим доступа: http://www.fao.org/faostat/en/#data/QC. Дата обращения: 16.01.2020.
- Беспалова Л.А., Кудряшов И.Н., Колесников Ф.А., Новиков А.В., Пузырная О.Ю., Грицай Т.И., Набоков Г.Д., Боровик А.Н., Керимов В.Р. Эволюция уборочного индекса и прогресс селекции зерновых культур на урожайность. Земледелие, 2014, № 3: 9-12.
- Силаева Л.П., Баринова Е.В. Современное состояние и условия рационального размещения производства пшеницы. Экономическийжурнал, 2019, 1(53): 33-42 (doi: 10.24411/2072-8220-2019-00003).
- Morgounov A., Zykin V., Belan I., Roseeva L., Zelenskiy Yu., Gomez-Becerrad H.F., Budakd H., Bekes F. Genetic gains for grain yield in high latitude spring wheat grown in Western Siberia in 1900-2008. Field Crops Research, 2010, 117: 101-112 (doi: 10.1016/j.fcr.2010.02.001).
- Плиско Л.Г., Пакуль В.Н. Адаптивная способность и стабильность сортов яровой мягкой пшеницы в условиях Западной Сибири. Международный научно-исследовательский журнал, 2016, 12(54, ч. 1): 140-144 (doi: 10.18454/IRJ.2016.54.228).
- Агеева Е.В., Лихенко И.Е., Советов В.В. Оценка экологической пластичности сортообразцов мягкой яровой пшеницы питомника Казахстанско-Сибирской сети CИММИТ. ДостижениянаукиитехникиАПК, 2018, 32(11): 26-29 (doi: 10.24411/0235-2451-2018-11106).
- Zhang L.Y., Liu D.C., Guo X.L., Yang W.L., Sun J.Z., Wang D.W., Zhang A. Genomic distribution of quantitative trait loci for yield and yield-related traits in common wheat. Journal of Integrative Plant Biology, 2010, 52(11): 996-1007 (doi: 10.1111/j.1744-7909.2010.00967.x).
- Sun C., Zhang F., Yan X., Zhang X., Dong Z., Cui D., Chen F. Genome-wide association study for 13 agronomic traits reveals distribution of superior alleles in bread wheat from the Yellow and Huai Valley of China. Plant Biotechnology Journal, 2017, 15: 953-969 (doi: 10.1111/pbi.12690).
- Yan X., Zhao L., Ren Y., Dong Z., Cui D., Chen F. Genome-wide association study revealed that the TaGW8 gene was associated with kernel size in Chinese bread wheat. Scientific Reports, 2019, 9: 2702 (doi: 10.1038/s41598-019-38570-2).
- Ain Q., Rasheed A., Anwar A., Mahmood T., Imtiaz M., Mahmood T., Xia X., He Z., Quraishi U.M. Genome-wide association for grain yield under rainfed conditions in historical wheat cultivars from Pakistan. Frontiers in Plant Science, 2015, 6: 743 (doi: 10.3389/fpls.2015.00743).
- Mwadzingeni L., Shimelis H., Rees D.J.G., Tsilo T.J. Genome-wide association analysis of agronomic traits in wheat under drought-stressed and non-stressed conditions. PLoS ONE, 2017, 12(2): e0171692 (doi: 10.1371/journal.pone.0171692).
- Liu C., Khodaee M., Lopes M.S., Sansaloni C., Dreisigacker S., Sukumaran S., Reynolds M. Multi-environment QTL analysis using an updated genetic map of a widely distributed Seri × Babax spring wheat population. Molecular Breeding, 2019, 39: 134 (doi: 10.1007/s11032-019-1040-1).
- Tshikunde N.M., Mashilo J., Shimelis H., Odindo A. Agronomic and physiological traits, and associated quantitative trait loci (QTL) affecting yield response in wheat (Triticum aestivum L.). A Review. Frontiers in Plant Science, 2019, 10: 1428 (doi: 10.3389/fpls.2019.01428).
- Entz M.H., Fowler D.B. Agronomic performance of winter versus spring wheat. Agronomy Journal, 1991, 83: 527-532 (doi: 10.2134/agronj1991.00021962008300030002x).
- Koppel R., Ingver A. A comparison of the yield and quality traits of winter and spring wheat. Latvian Journal of Agronomy, 2008, 11: 83-89.
- Крупнова О.В. О сопоставлении качества зерна яровой и озимой пшеницы в связи с делением на рыночные классы. Сельскохозяйственная биология, 2013, 48, 15-25 (doi: 10.15389/agrobiology.2013.1.15rus).
- Shoran J., Kant L., Singh R.P. Winter and spring wheat: an analysis of combining ability. Cereal Research Communication, 2003, 31: 347-354 (doi: 10.1007/BF03543364).
- Shah A.A., Mondal S.K., Rather I., Khurshid H., Wani A.A. Study of combining ability and gene action in F1 generation of winter × spring wheat derivatives. Journal of Pharmacognosy and Photochemistry, 2018, 7: 649-654.
- Grant M.N., McKenzie H. Heterosis in F1 hybrids between spring and winter wheats. Canadian Journal of Plant Science, 1970, 50: 137-140 (doi: 10.4141/cjps70-026).
- Kant L., Mani V.P., Gupta H.S. Winter × spring wheat hybridization — a promising avenue for yield enhancement. Plant Breeding, 2008, 120: 255-258 (doi: 10.1046/j.1439-0523.2001.00587.x).
- Мухордова М.Е. Изменчивость и генетический контроль зимостойкости у растений мягкой озимой пшеницы. Вестник НГАУ, 2019, 2: 35-41. (doi: 10.31677/2072-6724-2019-51-2-35-41).
- Коренюк Е.А., Мешкова Л.В. Комбинационная способность сортов и перспективных линий яровой мягкой пшеницы по признакам продуктивности в условиях южной лесостепи Омской области. Достижения науки и техники АПК, 2013, 5: 6-8.
- Косенко С.В. Комбинационная способность и генетический контроль продуктивной кустистости озимой мягкой пшеницы. Вестник АГАУ, 2017, 8: 6-9.
- Leonova I.N., Stasyuk A.I., Skolotneva E.S., Salina E.A. Enhancement of leaf rust resistance of Siberian winter wheat varieties by marker-assisted selection. CerealResearchCommunication, 2017, 45: 621-632 (doi: 10.1556/0806.45.2017.048).
- Стасюк А.И., Леонова И.Н., Салина Е.А. Проявление хозяйственно важных признаков у яровых гибридов мягкой пшеницы, отобранных с помощью MAS-технологии при скрещивании озимых сортов с яровыми донорами устойчивости к бурой ржавчине. Сельскохозяйственная биология, 2017, 52(3): 526-534 (doi: 10.15389/agrobiology.2017.3.526rus).
- Stasyuk A.I., Leonova I.N., Salina Е.А. Marker-assisted selection and yield component assessment of spring wheat plants derived from crosses between spring and winter wheat cultivars. Proc. of the 5th Int. Sci. Conf. PlantGen, «Current Challenges in Plant Genetics, Genomics, Bioinformatics, and Biotechnology» /A. Kochetov, E. Salina (eds.). Novosibirsk, 2019: 193-195 (doi: 10.18699/ICG-PlantGen2019-62).
- Song Q.J., Shi J.R., Singh S., Fickus E.W., Costa J.M., Lewis J., Gill B.S., Ward R., Cregan P.B. Development and mapping of microsatellite (SSR) markers in wheat. Theoretical and Applied Genetics, 2005, 110: 550-560 (doi: 10.1007/s00122-004-1871-x).
- Удачин Р.А., Головоченко А.П. Методика оценки экологической пластичности сортов пшеницы. Селекция и семеноводство, 1990, 5: 2-6.
- Доспехов Б.А. Методика полевого опыта (с основами статистической обработки результатов исследований). М., 1973.
- Hammer O., Harper D.A.T., Ryan P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontologia Electronica, 2001, 4(1): 1-9.
- Пискарев В.В., Тимофеев А.А., Цильке Р.А. Результаты селекции яровой мягкой пшеницы на продуктивность в условиях лесостепи западной Сибири. Сибирский вестник сельскохозяйственной науки, 2010, 8: 18-24.
- Василова Н.З., Асхадуллин Данил Ф., Асхадуллин Дамир Ф., Багавиева Э.З., Тазутдинова М.Р., Хусаинова И.И., Насихова Г.Р. Влияние условий выращивания на формирование урожайности яровой мягкой пшеницы. Достижения науки и техники АПК, 2015, 29(11): 41-43.
- Валекжанин В.С., Коробейников Н.И. Адаптивные реакции сортов и линий яровой мягкой пшеницы на агроэкологические условия в Алтайском крае. Сибирский вестник сельскохозяйственной науки, 2008, 3(183): 31-36.
- Gonzalez-Navarro O.E., Griffithsa S., Molero G., Reynolds M.P., Slafer G.A. Variation in developmental patterns among elite wheat lines and relationships with yield, yield components and spike fertility. Field Crops Research, 2016, 196: 294-304 (doi: 10.1016/j.fcr.2016.07.019).
- Агеева Е.В., Лихенко И.Е., Советов В.В., Пискарев В.В. Экологическая пластичность пшеницы в лесостепи Западной Сибири. Вестник НГАУ, 2015, 1(34): 22-28.
- Артемова Г.В., Степочкин П.И., Пономаренко В.И., Ермошкина Н.Н., Пономаренко Г.В. Оценка сортов озимой пшеницы Сибирской селекции по параметрам экологической пластичности и стабильности. Сибирский вестник сельскохозяйственной науки, 2015, 6(247): 5-11.
- Волкова Л.В. Оценка сортов яровой мягкой пшеницы по урожайности и адаптивным свойствам. Аграрная наука Евро-Северо-Востока, 2017, 4(59): 19-23.
- Кадычегов А.Н., Бородыня А.Н., Кадычегова В.И. Влияние предшественника на экологическую пластичность и стабильность урожайности сортов яровой мягкой пшеницы различных групп спелости в степной зоне юга средней Сибири. Вестник АГАУ, 2015, 7 (129): 11-15.
- Slafer G.A., Savin R., Sadras V.O. Coarse and fine regulation of wheat yield components in response to genotype and environment. FieldCropsResearch, 2014, 157: 71-83 (doi: 10.1016/j.fcr.2013.12.004).
- Пушкарев Д.В., Чурсин А.С., Кузьмин О.Г., Краснова Ю.С., Каракоз И.И., Шаманин В.П. Экологическая пластичность и стабильность сортов яровой мягкой пшеницы в степной зоне Омской области. Вестник ОмГАУ, 2017, 4(28): 61-67.
- Eberhart S.A., Russell W.A. Stability parameters for comparing varieties. Crop Science, 1966, 6(1): 36-40 (doi: 10.2135/cropsci1966.0011183X000600010011x).
- Tai G.C.C. Genotypic stability analysis and application to potato regional trials. Crop Science, 1971, 11(2): 184-190 (doi: 10.2135/cropsci1971.0011183X001100020006x).
- Потанин В.Г., Алейников А.Ф., Стёпочкин П.И. Новый подход к оценке экологической пластичности сортов растений. Вавиловский журнал генетики и селекции, 2014, 18(3): 548-552.
- Куркова И.В., Терехин М.В. Оценка экологической пластичности сортов яровой мягкой пшеницы селекции Дальнего Востока. Вестник АГАУ, 2008, 7(45): 8-11.
- Friebe B., Jiang J., Raupp W.J., McIntosh R.A., Gill B.S. Characterization of wheat-alien translocation conferring resistance to diseases and pests: current status. Euphytica, 1996, 91: 59-87 (doi: 10.1007/BF00035277).
- Hysing S.C., Hsam S.L.K., Singh R.P., Huerta-Espino J., Boyd L.A., Koebner R.M.D., Cambron S., Johnson J.W., Bland D.E., Liljeroth E., Merker A. Agronomic performance and multiple disease resistance in T2BS.2RL wheat-rye translocation lines. Crop Science, 2007, 47: 254-260 (doi: 10.2135/cropsci2006.04.0269).
- Leonova I.N., Budashkina E.B. The study of agronomical traits determining the productivity of the Triticum aestivum/Triticum timopheevii introgression lines with resistance to fungal diseases. Russian Journal of Genetics: Applied Research, 2017, 7(3): 299-307 (doi: 10.1134/S2079059717030091).
- Крупнов В.А., Сибикеев С.Н. Чужеродные гены для улучшения мягкой пшеницы. Идентифицированный генофонд растений и селекция /Под ред. Б.В. Ригина, Е.И. Гаевской. СПБ, 2005: 740-758.