








УДК 632.7:632.08
ПРОСТРАНСТВЕННОЕ РАСПРЕДЕЛЕНИЕ ЖУЖЕЛИЦ И СТАФИЛИНИД В АГРОЭКОСИСТЕМЕ
О.Г. ГУСЕВА, А.Г. КОВАЛЬ
Провели сравнительное изучение видового состава, динамической плотности и структуры доминирования многоядных хищных жуков (жужелиц и стафилинид) на участках агроэкосистемы с различными экологическими условиями. Открытые участки заселяли преимущественно мелкие светолюбивые виды жесткокрылых, поля многолетних трав и обочины — тенелюбивые виды этих насекомых. Поля многолетних трав служили источниками обогащения видового состава жуков на других участках агроэкосистемы, особенно на полях озимых и яровых зерновых культур.
Ключевые слова: жужелицы, стафилиниды, многолетние травы, агроэкосистема.
Энтомофаги представляют собой один из основных элементов биоценотической саморегуляции в агроэкосистемах. В зависимости от выращиваемой культуры и системы обработки почвы на разных участках формируются неодинаковые условия микроклимата, освещенности, скважности и т.д. Растительный покров уменьшает суточное колебание температуры, изменяет режим теплообеспеченности и освещенности, увеличивает влажность воздуха (1). Смена растительного покрова сопровождается значительными модификациями структуры комплекса членистоногих, обитающих на поверхности почвы (2).
В настоящее время накоплено много информации о видовом составе напочвенных жуков (Insecta, Coleoptera), среди которых выделяются жужелицы (Carabidae, карабиды) и стафилиниды (Staphylinidae, стафилины) — наиболее многочисленные многоядные хищники (3-5). Однако в большинстве случаев эти данные получены в удаленных друг от друга географических зонах, на полях, занятых разными культурами, и потому почти не сопоставимы (6).
Целью нашей работы была оценка степени пригодности различных полей и окружающих биотопов для хищных напочвенных жесткокрылых, в связи с чем мы провели сравнительное изучение видового состава, динамической плотности и структуры доминирования жужелиц и стафилинид на участках агроэкосистемы с различными экологическими условиями.
Методика. Исследования выполняли в 2004-2006 годах на территории Меньковской опытной станции Агрофизического НИИ (Гатчинский р-н, Ленинградская обл.). Агроэкосистема включала отдельные поля зерно-тра-вяно-пропашного севооборота (чистого пара, озимых зерновых, картофеля, клевера с подсевом тимофеевки и вики, яровых зерновых с подсевом многолетних трав) и окружающие территории между возделываемыми землями и лесным массивом, заросшие разнотравьем и кустарниками. Площадь одного поля — 0,6 га, обследованных участков — около 5 га.
В качестве ловушек для напочвенных жуков использовали стеклянные банки объемом 0,5 л с диаметром отверстия 72 мм, заполненные на 1/3-1/2 4 % раствором формалина (7). В центральной части каждого экспериментального поля устанавливали по 10 ловушек на расстоянии 10 м одна от другой. На обочине ловушки располагали в ряд параллельно краю последнего поля севооборота на расстоянии 10 м друг от друга и 2 м от границы обрабатываемого участка. Учеты проводили с мая по август. Попадание жужелиц и стафилинид в ловушки (уловистость) отражало динамическую плотность этих жуков. Полученные по ловушкам данные пересчитывали на единицу уловистости — 10 ловушко-суток (10 л.-с.) (8).
В качестве показателя видового богатства использовали индекс Маргалефа (Dmg): Dmg = (S - 1)/lnN, где S — число выявленных видов, N — общее число особей всех видов. Вариабельность средних значений уловистости карабид и стафилинов характеризовалась величиной ошибки средней для 0,5 % уровня значимости (р <= 0,05). При оценке взаимного обогащения комплексов напочвенных хищных жуков, складывающихся в агроэкосистеме, анализировали видовые списки жужелиц и стафилинид отдельных полей севооборота. Для списков, представленных в виде семейства множеств, проводили расчет мер включения и составляли матрицу мер включения (9). Если видовой список одного поля полностью входит в список другого, то его включение считают равным 100 %, и первое поле характеризуется как банальное. Сопоставление мер включения возможных попарных сочетаний позволяло выяснить закономерности, отражающие взаимоотношения комплексов хищников отдельных полей севооборота.
Результаты. За годы исследований в изучаемой агроэкосистеме было собрано свыше 13 тыс. особей жужелиц (69 видов) и 3 тыс. — стафилинид (69 видов). Наибольшее видовое богатство жесткокрылых отмечали на участках, окружающих поля. Показатели Dmg в этом биотопе для жужелиц и стафилинид составили соответственно 5,4 и 5,5 (табл.). На затененных обочинах встречались некоторые лесные виды, никогда не регистрировавшиеся на полях (жужелицы Carabus hortensis L., Badister lacertosus Sturm, Calathus micropterus Duft., стафилины Staphylinus erythropterus L., Oxypoda abdominalis Mnnh., Ocalea badia Er. и др.). В то же время среди густого травостоя и кустарников отсутствовали многие виды карабид из рода Bembidion, предпочитающие открытые освещенные участки.
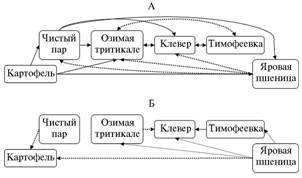 |
Рис. 1. Связь между комплексами напочвенных хищников — жужулиц (А) и стафилинид (Б) на разных полях севооборота. Сплошные и пунктирные линии — орграфы отношений банальности В75 и В70 (включение списка видов одного поля в список другого соответственно не менее 75 и 70 %). Число входящих стрелок отражает разнообразие комплекса. |
В севообороте самые высокие показатели видового разнообразия жу-желиц были отмечены на полях яровых зерновых с подсевом многолетних трав, а также клевера с подсевом тимофеевки и вики, стафилинид — на полях картофеля (рыхлая почва гребней привлекательна для видов, передвигающихся в почвенных скважинах), клевера с подсевом тимофеевки и вики, а также ози-мой тритикале (см. табл.). Поля многолетних трав, особенно клевера с подсевом тимофеевки, были наименее банальными, то есть наиболее богатыми по видовому составу, особенно в первую половину вегетации. Они служили источниками увеличения видового разнообразия фаунистических комплексов других полей, преимущественно озимых и яровых зерновых культур (рис. 1).
Поле чистого пара было наиболее бедным по видовому составу жужелиц и стафилинид (соответственно Dmg = 3,8 и Dmg = 2,8), однако оно оказалось благоприятным местом обитания для мелких представителей подсемейства Aleocharinae и рода Bembidion. Последние (особенно B. quadrima-culatum, B. properans и B. lampros)предпочитают освещенные участки с редкой растительностью (10). В первую половину сезона они также преобладали на полях картофеля и яровой пшеницы с подсевом тимофеевки (см. табл., рис. 2). Очевидно, в фазу всходов микроклимат таких участков и чистого пара практически не различался (1). В июле, когда вегетативная масса культурных растений значительно увеличивалась, на полях яровых зерновых и картофеля наблюдалось резкое снижение числа карабид из указанного рода. В то же время на соседнем поле, занятом озимой тритикале, доля жужелиц рода Bembidion в мае составила 8 %, а в дальнейшем на этом поле отмечались только отдельные их особи.
| Видовое богатство, динамическая плотность и структура доминирования хищных напочвенных жуков на разных участках агроэкосистемы (Ленинградская обл., 2004-2006 годы) | ||||||
Участок, |
Carabidae (жужелицы) |
Staphylinidae (стафилиниды) |
||||
А |
Б |
В |
А |
Б |
В |
|
Чистый пар |
3,8 |
48,7±12,96 |
Bemdidionquadrimacula- |
2,8 |
5,9±4,61 |
Aleochara bipustulata L. (33,8) |
Картофель |
4,5 |
13,8±2,61 |
B. properans (16,1) |
4,8 |
5,6±3,29 |
A. bipustulata (19,8) |
Вико-овсяная смесь |
4,7 |
27,1±4,48 |
P. cupreus (20,6) |
3,2 |
5,7±0,64 |
Philonthus cognatus |
Яровая пшеница с подсевом тимофеевки |
5,3 |
14,8±2,36 |
B. quadrimaculatum (19,3) |
3,4 |
5,6±1,68 |
D. angustula (31,0) |
Клевер с подсевом тимофеевки и вики |
5,1 |
41,9±5,91 |
P. cupreus (59,3) |
4,6 |
2,0±0,75 |
Ph. cognatus (29,8) |
Озимая тритикале |
4,8 |
40,3±10,35 |
P. cupreus (51,2) |
4,5 |
5,0±1,01 |
Ph. cognatus (17,4) |
Обочины полей |
5,4 |
9,8±2,11 |
Poecilus versicolor Sturm (51,2) |
5,5 |
13,9±3,51 |
Drusilla canaliculata F. (73,8) |
П р и м е ч а н и е. А — видовое богатство, Dmg; Б — динамическая плотность, число особей на 10 ловушко-суток (M±m); В — доминирующие виды (доля от общего числа особей, %). |
||||||
Более крупные виды жужелиц и стафилинид населяли в основном затененные участки. Подобный факт также отмечали, например, в условиях Чехии (2). Жужелицы Harpalusrufipes и Poeciluscupreus предпочитали поля, занятые озимыми зерновыми культурами и многолетними травами. Так, на участке с озимой тритикале к виду P. cupreus относилось 51,2 % особей карабид, средняя длина тела которых составляла 12,1 мм. На поле, занятом чистым паром, отмечена наибольшая доля особей B. quadrimaculatum, имевших средний размер 3,2 мм. На полях яровых зерновых в зависимости от изреженности посевов преобладали виды из рода Bembidion, а также P. cupreusи H. rufipes (см. табл.).
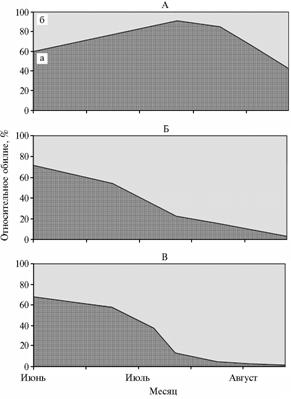 |
Рис. 2. Сезонные изменения доли жужелиц из рода Bembidion (а) и остальных карабид (б) на полях севооборота под различными культурами: А — чистый пар, Б — яровая пшеница, В — картофель (Ленинградская обл., 2004-2006 годы). |
Аналогичная ситуация наблюдалась и при распределении в агробиоценозе различных видов стафилинид. Так, для освещенных участков были характерны высокие показатели относительного обилия мелких видов — представителей подсемейства Aleocharinae (Aloconota gregaria, Dinaraea angustula, Aleochara bipustulata), длина тела которых составляла в среднем 3,2-3,3 мм (см. табл.). На полях, занятых вико-ов-сяной смесью и яровой пшеницей с подсевом тимофеевки, эти виды преобладали в июне, а на участке, занятом чистым паром, — в течение всего сезона. После увеличения вегетативной массы растений повышалась численность крупного тенелюбивого стафилина Philonthus cognatus. На поле с вико-овсяной смесью с густым травостоем доля особей этого вида достигала 33,1 %, на поле яровой пшеницы с подсевом тимофеевки — 7,8 %. На полях, занятых озимой тритикале, а также клевером с подсевом тимофеевки и вики, Ph. cognatus был самым многочисленным видом, особенно в первую половину сезона. На участке с озимой тритикале доля стафилинид из рода Philonthus размером 9,5 мм составляла 33,5 % (наиболее высокий показатель среди всех размерных групп стафилинид). Возможная причина заключается в том, что большинство крупных стафилинид предпочитают 100 % влажность воздуха, а средние и мелкие формы весьма сухоустойчивы (11).
Следует отметить, что при значительных различиях в структуре доминирования стафилинид на многих полях получены близкие значения средней за сезон динамической плотности этих жесткокрылых. На поле тимофеевки с клевером второго года пользования показатель был наименьшим — 2,0 экз. на 10 л.-с. (см. табл.), на других участках он колебался от 5,0 до 5,9 экз. на 10 л.-с. В целом динамическая плотность стафилинид оказалась значительно ниже, чем жужелиц (см. табл.). На необрабатываемых обочинах полей, напротив, выявили наименьшую в агроэкосистеме динамическую плотность жужелиц (9,7 экз. на 10 л.-с.) и наибольшую — стафилинов (13,8 экз. на 10 л.-с.) (см. табл.). Следовательно, обрабатываемые земли более благоприятны для обитания многих видов жужелиц.
Среди жужелиц, собранных на обочинах, 51,2 % особей относились к виду Poecilus versicolor. На полях севооборота этот вид был немногочисленным, причем наибольшее число представителей P. versicolor отмечали на участках с многолетними травами. Доля другого близкого вида — P. cupreusна обочинах составила 11,6 %. Известно, что обочины служат местом зимовки для жужелиц P. cupreus(12). Более 15 % от общего числа карабид, собранных на обочинах, приходилось на лесные виды Pterostichusniger и P. oblongopunctatus. На полях с густым травостоем регистрировали только отдельных особей этих видов, а на открытых участках они не встречались.
Среди стафилинид, обитающих на обочине, доминировали редкие для обрабатываемых полей виды. Так, в 2006 году 73,8 % особей, собранных в этом биотопе, относились к виду Drusilla canaliculata, 7,8 % — к Sepedophilus marschami. Исследования, проведенные в Московской области, также подтвердили, что D. сanaliculata всегда предпочитает обочины полям и встречается только в краевой зоне последних (13). Работы, выполненные в Чехии, показали сходные результаты: особи этого вида чаще обитают на затененных, заросших сорняками участках полей (2).
Таким образом, условия, складывающиеся на отдельных участках в агроэкосистеме, способствуют формированию специфических комплексов, различающихся по видовому составу, динамической плотности и структуре доминирования напочвенных хищных жуков. Открытые пространства заселяют преимущественно мелкие светолюбивые виды жужелиц и стафилинид, поля многолетних трав и обочины — более крупные тенелюбивые виды этих жесткокрылых. Поля многолетних трав, особенно в первую половину вегетации, служат источниками обогащения видового состава жуков на других участках в агроэкосистеме и, прежде всего, на полях яровых и озимых зерновых культур. Обочины характеризуются высокими показателями видового богатства за счет присутствия лесных видов, большая часть которых на полях редки или вообще не встречаются.
Л и т е р а т у р а
1. С а п о ж н и к о в а С.А. Микроклимат и местный климат. Л., 1950.
2. H o n e k A. The effect of plant cover and weather on the activity density of ground surface arthropods in a fallow field. Biological Agriculture and Horticulture (Cambridge, UK), 1997, 15: 203-210.
3. С о б о л е в а - Д о к у ч а е в а И.И., С о л д а т о в а Т.А. Влияние экологических условий сельскохозяйственной культуры на хищных почвенных жесткокрылых (Coleoptera, Carabidae, Staphylinidae). В сб.: Фауна и экология почвенных беспозвоночных Московской области. М., 1983: 120-130.
4. Ш а р о в а И.Х. Жизненные формы жужелиц (Coleoptera, Carabidae). М., 1981.
5. The agroecology of carabid beetles /J.M. Holland (ed.). Andover, UK, 2002.
6. Б е л о у с о в И.А. Факторы, определяющие карабидокомплексы в агроценозах и пути их обогащения. В сб. науч. тр. Всесоюз. НИИ защиты растений: Интродукция, акклиматизация и селекция энтомофагов. Л., 1987: 55-64.
7. H e y d e m a n n B. Über die Bedeutung der «Formalinfallen» für die zoologische Landesforschung. Faunistische Mitteilungen aus Norddeutschand (Kiel, Deutschland), 1956, 6: 19-24.
8. Ш а р о в а И.Х. Особенности биотопического распределения жужелиц (Coleoptera, Ca-rabidae) в зоне смешанных лесов Подмосковья. Уч. зап. Московского гос. пед. ин-та им. В.И. Ленина, 1974, 465: 61-86.
9. А н д р е е в В.Л. Классификационные построения в экологии и систематике. М., 1980.
10. L i n d r o t h C.H. The Carabidae (Coleoptera) of Fennoscandia and Denmark. Leiden, Netherlands, Copenhagen, Denmark, 1985.
11. Т и х о м и р о в а А.Л. Морфоэкологические особенности и филогенез стафилинид (с каталогом фауны СССР). М., 1973.
12. Т и м о х о в а О.В. Пространственно-временное размещение жужелиц (Coleoptera, Ca-rabidae) в агроэкосистеме озимой пшеницы. Автореф. канд. дис. М., 2001.
13. С о б о л е в а - Д о к у ч а е в а И.И., Ч е р н ы ш е в В.Б.,
А ф о н и н а В.М., Т и
м о х о в А.В. Сезонная динамика размещения массовых видов стафилинид (Coleoptera, Staphylinidae) в агроэкосистемах. Зоол. журн., 2002, 81(4): 451-456.
REGIONAL DISTRIBUTION OF Carabidae AND Staphylinidae IN AGROECOSYSTEM
O.G. Guseva, A.G. Koval’
The authors made a comparative study of species content, dynamic density and domination structure in plurivorous rapacious beetles (Carabidae and Staphylinidae) on areas of agroecosystem with various ecological conditions. The small light-requiring species occupied as rule the exposed areas, shade-requiring coleopteran — the fields with permanent grasses and waysides. The fields with permanent grasses are source of enrichment of species composition of beetles, inhabited in other areas, areas with winter and spring seed crops, particularly.
Keywords: carabid beetles, staphylinid beetles, perennial grasses, agroecosystem.
ФГОУ ВПО Ульяновский государственный |
Поступила в редакцию |




