








УДК 633.14:631.52:631.527.5
СЕЛЕКЦИЯ ОЗИМОЙ РЖИ ПО МЕТОДУ ПАРНЫХ СКРЕЩИВАНИЙ
А.А. ГОНЧАРЕНКО
Предложена схема селекционного процесса озимой ржи с использованием метода парных скрещиваний. Анализируются селекционные преимущества новой схемы, дается расчет уровня инбридинга в популяциях ржи, полученных при объединении изолированно размноженных полусибсовых и полносибсовых потомств. Показано, что возникающий инбридинг можно снизить до нулевого уровня, используя свободное переопыление соответствующего числа семей. Минимально допустимое число родительских компонентов при парных скрещиваниях должно составлять около 40.
Ключевые слова: озимая рожь, метод резервов, парные скрещивания, полные сибсы, коэффициент инбридинга.
В настоящее время производство озимой ржи в России переживает глубокий кризис: посевные площади и валовые сборы зерна уже сократились более чем 3-кратно, и эта тенденция все еще сохраняется, что обусловлено прежде всего низкой конкурентоспособностью ржи по сравнению с другими культурами. Так, рост урожайности ржи не происходит такими же относительно высокими темпами, как, например, по главной зерновой культуре — озимой пшенице. За последние 50 лет прирост урожайности ржи в Российской Федерации составил 16,5, по озимой пшенице — 26,7 кг в год, или в 1,6 раза больше. Возделываемые сорта ржи относительно слабо реагируют на повышение культуры земледелия и применение интенсивных технологий. Причина кроется в слабой селекционной проработке возделываемых сортов по ряду признаков, и прежде всего по устойчивости к полеганию, а если смотреть дальше — в методическом несовершенстве схем и методов селекции, на основе которых они созданы.
Рожь как строго перекрестно опыляемая культура с малым коэффициентом размножения является трудным объектом для селекции (1). Широко применяемый массовый отбор растений по фенотипу на фоне свободного переопыления не всегда обеспечивает желаемые результаты, особенно по признакам с низкой наследуемостью. Поэтому неудивительно, что технология селекции этой культуры изобилует большим числом вариаций и комбинаций различных схем и методов отбора.
Большинство отечественных сортов ржи создано методом многократного индивидуально-семейного отбора из гибридных популяций. Указанный метод не предполагает строгого сохранения первоначально отобранных полусибсовых потомств (семей), худшие из которых выбраковываются, в основном после свободного переопыления. В этом случае известен генотип только материнского растения семьи, а генотип отцовских форм неизвестен, что исключает возможность их выбраковки. Поэтому более правильно было бы такой вариант отбора определить как наполовину индивидуальный и наполовину массовый. Подобная половинчатость в принципе нежелательна, поскольку снижается результативность отбора (2).
Более эффективен в этом отношении семейный отбор по методу резервов (половинок). У ржи его впервые применил В. Лаубе в 1921 году в Германии (3). Как более совершенный вариант семейного отбора он позволяет более направленно управлять процессом опыления и получать улучшенную популяцию. Однако и в этом случае при свободном переопылении родословная отобранных потомств по отцовской линии контролируется лишь частично. Стремление к более полному контролю породило субвариант описанного метода с изолированным размножением семей под изоляторами после проведения первого цикла отбора.
Описание методики. В последние годы широкое применение в селекции ржи получил метод, в котором объектом отбора служат не полусибсовые, а полносибсовые (полносестринские) семьи (нем. Vollgeschwisterfamilien — VGF). Впервые замену полусибсовых семей на полносибсовые использовал T. Wolski в Польше (1). При создании сорта Даньковске злоте он параллельно с отбором полусибсов проводил парные скрещивания предварительно отобранных растений. В основу этого методического приема он положил концепцию R.V. Sengbusсh (4), предложившего во избежание инбридинга переопылять растения из разных семей. Суть способа состоит во взаимном переопылении двух самонесовместимых растений (то есть пары) под пергаментным изолятором и последующем испытании полученного потомства по методу резервов. Схематично процедура такого селекционного процесса показана на рисунке 1.
В 1-й год закладывается питомник парных скрещиваний, где семена селектируемой популяции высеваются ленточным способом по схеме примерно 20x20x50 см. В период колошения в питомнике проводят парные скрещивания визуально лучших растений. На два растения из разных семей перед цветением надевают пергаментный изолятор, под которым происходит их взаимное переопыление. Переопыляемые растения высокосамонесовместимы, а потому имеют низкую степень самоопыления и высокую — перекрестного опыления, то есть образуют гибридные семена, что очень желательно для дальнейшей селекции. Потомство такого гибрида не разделяют в зависимости от того, какое растение было материнским, а какое отцовским. Поэтому семена от растений каждой пары смешивают, и они представляют собой как бы потомство одного растения. Объем парных скрещиваний должен быть большим (~ 5000 пар), с тем чтобы обеспечить интенсивный (10-20 %) отбор потомств в последующие годы. Убранные пары обмолачивают каждую отдельно и отбирают те, у которых образовалось не менее 300 зерен.
На 2-й год закладывают сравнительное микроиспытание А-семей без повторений со стандартом через несколько делянок. Для посева используют только часть (100-150) семян изучаемых семей, а оставшиеся сохраняют в резерве. По сути, прием представляет собой временную изоляцию полносибсовых потомств, позволяющую исключить худшие семьи из дальнейшего переопыления с лучшими. Для последующего посева используют только те семьи, которые при полевых испытаниях получили высокие оценки.
На 3-й год в условиях экранной (стенки типа «шпанвенде») или более строгой (тканевой, пленочной) изоляции высевают резервные семена только тех потомств, которые по результатам сравнительного испытания оказались лучшими. Происходит внутрисемейное переопыление, и полученных семян образуется достаточно для того, чтобы на 4-й год, сохранив резерв семян, провести широкое экологическое испытание семей в 3-4 пунктах в трех повторениях с применением сеялочного посева и производственной нормой высева. По результатам экологического испытания отбирают примерно 40 лучших А1-семей и, используя резервы, проводят их переопыление. В итоге получают генетически улучшенную популяцию. Одновременно часть резервных семян отобранных семей используют для проведения второго цикла парных скрещиваний.
Основная трудность в реализации приведенной схемы состоит в получении большого числа полносибсовых А-семей и изолированном размножении лучших из них. Однако затраты стоят того, так как селекционер получает возможность полностью контролировать генотип обоих родителей, что повышает эффективность и ускоряет отбор. В этом случае потомство двух взаимно переопыленных растений представляет собой последний вариант, сохраняющий нужные признаки, который может использовать селекционер, не прибегая к инбридингу.
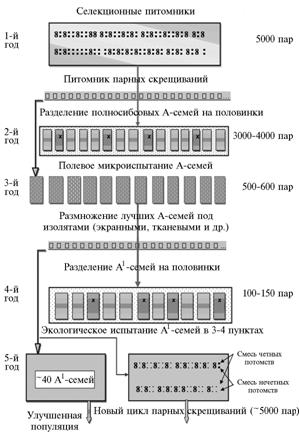 |
Рис. 1. Схема селекции озимой ржи с использованием метода резервов и парных скрещиваний. |
Описанная выше процедура представляет собой отбор полных сибсов с применением изолированного размножения потомств. Обычно на практике селекционеры стараются этого избегать, чтобы не допустить инбридинга, и такая позиция обоснованна. Однако в этом есть и свои плюсы, особенно если учесть, что возникший инбридинг можно устранить и довести до нулевого уровня посредством переопыления соответствующего числа семей. Выгода же состоит в том, что при изолированном размножении генетически ценные свойства материнского и отцовского родителей легко фиксируются и сохраняются, а от нежелательных можно избавиться последующей выбраковкой плохих потомств.
Естественно, переопыление в пределах семьи приводит к родственному скрещиванию, что неблагоприятно отражается на жизнеспособности и урожайности популяции. Проблема усугубляется, если в селектируемую популяцию объединяется малое число родоначальных семей. В этом случае возникает дополнительный инбридинг, который еще больше снижает продуктивность популяции.
Концепция о необходимости сохранения максимальной гетерозиготности популяций ржи для достижения высокой продуктивности общепризнана. H. Nilsson (5) и A. Muntzing (6) определили рожь как сильно чувствительную к инбридингу. Из селекционной практики также известно, что при проведении отбора не следует недооценивать возможность депрессии от возникающего инбридинга в популяциях ржи (7-9).
Известно, что инбредная депрессия уменьшается, если число родителей увеличивается. Однако в этом случае средняя продуктивность популяции повышается только до определенного предела, затем рост прекращается. Объясняется это нивелирующим эффектом возрастающего числа родительских компонентов. Каждая новая родительская форма, добавленная к уже отобранным, хотя и уменьшает потери, обусловленные инбредной депрессией, все же мало способствует увеличению продуктивности популяции из-за ее недостаточно высокой комбинационной способности. Подтверждением служит тот факт, что с увеличением числа отбираемых семей закономерно снижается селекционный дифференциал, а селектируемая популяция приближается к исходному уровню.
Согласно T.H. Busbice (10), инбридинг в синтетических популяциях обратно пропорционален числу (n) переопыляемых родительских генотипов (F = 1/2n). Однако при использовании полусибсов и полных сибсов функциональная зависимость между F и n несколько видоизменяется, так как в этом случае число родителей определяется не только числом переопыляемых семей, но и числом генотипов внутри каждой семьи, коэффициент родства между которыми равен 0,25 в случае полусибсов и 0,50 в случае полных сибсов. Последнее может вносить существенную поправку в уровень инбридинга популяции, поскольку при размножении семьи под изоляторами ее численность увеличивается. При этом вероятность внутрисемейного скрещивания родственных генотипов заметно возрастает, что влечет дополнительное повышение уровня инбридинга.
| 1. Вероятный коэффициент инбридинга (F) в популяциях ржи, полученных при переопылении различного числа полусибсовых и полносибсовых семей | ||||
Число переопыляемых семей (n) |
Полусибсы |
Полные сибсы |
||
свободно переопыленные |
изолированно размноженные |
свободно переопыленные |
изолированно размноженные |
|
1 |
0,1250 |
0,1406 |
0,2500 |
0,3125 |
5 |
0,0250 |
0,0281 |
0,0500 |
0,0625 |
10 |
0,0125 |
0,0141 |
0,0250 |
0,0312 |
15 |
0,0083 |
0,0094 |
0,0167 |
0,0208 |
20 |
0,0062 |
0,0070 |
0,0125 |
0,0156 |
25 |
0,0050 |
0,0056 |
0,0100 |
0,0125 |
30 |
0,0042 |
0,0047 |
0,0083 |
0,0104 |
35 |
0,0036 |
0,0040 |
0,0071 |
0,0089 |
40 |
0,0031 |
0,0035 |
0,0062 |
0,0078 |
45 |
0,0028 |
0,0031 |
0,0056 |
0,0069 |
50 |
0,0025 |
0,0028 |
0,0050 |
0,0062 |
Сравнительную оценку коэффициента инбридинга в популяциях ржи, полученных при отборе полусибсовых и полносибсовых потомств, мы провели на основе уравнений, предложенных нами ранее (11). Полученные результаты (табл. 1, рис. 2) свидетельствуют, что в обоих вариантах отбора при увеличении числа родоначальных семей уровень инбридинга популяций закономерно снижался и только при переопылении 35-40 семей и более его величина составляла менее 1 %. Особенность состоит в том, что в случае изолированного размножения полносибсовых семей уровень инбридинга синтезируемых популяций при одинаковом числе родительских компонентов в 2,22 раза выше, чем при отборе полусибсовых. Причем это превышение сохраняется при любом числе родоначальных семей. Если вместо изолированного размножения семей применить их свободное переопыление, то различие в уровне инбридинга между сравниваемыми вариантами будет 2-кратным, что объясняется 2-кратной разницей в степени родства между полусибсами и полными сибсами. Добавочный инбридинг (0,22 раза) обусловлен тем, что при синтезе улучшенной популяции в переопылении участвовали уже инбредные А1-семьи, у которых коэффициент инбридинга (F) составил 0,25 в результате изолированного размножения.
Ранее мы показали (12), что при возрастании коэффициента инбридинга на 1,0 % урожайность популяций ржи снижается на 1,2 %. Если исходить из этой пропорции, то в случае отбора полусибсов в минимуме можно ограничиться переопылением 20 лучших А1-потомств, так как вероятный коэффициент инбридинга (F) составит 0,70 %, а снижение урожайности от инбредной депрессии не превысит 0,84 %. В случае парных скрещиваний и отбора изолированно размноженных полносибсовых потомств аналогичный уровень инбридинга селектируемая популяция будет иметь при переопылении 40 А1-потомств (F = 0,78 %), то есть при их удвоенном числе. Мы полагаем, что это число родительских компонентов можно считать минимально допустимым при отборе полносибсовых потомств.
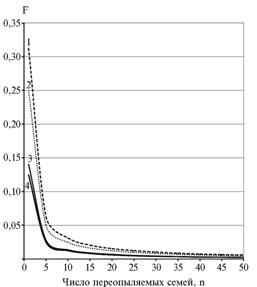 |
Рис. 2. Коэффициент инбридинга (F) в популяциях ржи при переопылении различного числа полусибсовых и полносибсовых семей: 1-2 — соответственно полные сибсы А1 и А; 3-4 — соответственно полусибсы А1 и А. |
Важно отметить, что при синтезе улучшенной популяции резервные семена лучших 40 А1-потомств не следует просто смешивать и свободно переопылять. Для того чтобы избежать дополнительного инбридинга и получить более выравненную популяцию, необходимо разделить отобранные потомства на две равные группы (например, по четным и нечетным номерам), затем семена в пределах группы смешать и высеять параллельными рядами для проведения межгрупповых парных скрещиваний. В этом случае потомство полученных пар будет неинбредным, что очень важно для синтеза улучшенной популяции. Такой прием позволяет снять инбридинг, обусловленный изолированным размножением полносибсовых семей. Что касается потенциального инбридинга в синтезируемой популяции, то он будет полностью определяться числом полносибсовых потомств, только уже не изолированно размноженных, а свободно переопыленных.
Экспериментальную проверку эффективности отбора потомств от парных скрещиваний мы провели в 2005-2008 годах. На базе 4 гибридных популяций были отобраны полусибсовые и полносибсовые потомства, полевое испытание которых выполняли в селекционном питомнике в один и тот же год (табл. 2). Основной вывод из этого сравнения сводится к тому, что в количественном отношении отбор полносибсовых семей на 10 % превосходит результаты, полученные при отборе полусибсов. О качестве отбираемых потомств можно судить по селекционной характеристике сорта озимой ржи Московская 12, полученного методом парных скрещиваний (табл. 3). Как видно из представленных данных, при отборе полносибсовых потомств можно улучшить популяцию по урожайности, зимостойкости, короткостебельности, устойчивости к полеганию, а также сформировать хорошо выравненный стеблестой.
| 2. Результаты отбора полусибсовых и полносибсовых А-потомств озимой ржи по методу резервов | |||||||
Селектируемая популяция |
Год оценки |
Полусибсовое потомство |
Полносибсовое потомство |
||||
изучено |
отобрано |
изучено |
отобрано |
||||
всего |
% |
всего |
% |
||||
ГК-870/99 |
2005 |
472 |
55 |
11,6 |
704 |
169 |
24,0 |
П-12/92 |
2006 |
313 |
67 |
21,4 |
552 |
168 |
30,4 |
П-11/92 |
2007 |
370 |
58 |
15,7 |
1052 |
301 |
28,6 |
ГК-984/01 |
2008 |
332 |
53 |
16,0 |
537 |
118 |
22,0 |
Среднее |
|
– |
– |
16,2 |
– |
– |
26,3 |
3. Селекционная характеристика сорта озимой ржи Московская 12, полученного методом парных скрещиваний (Московская обл., 2008 год) |
|||||||
Сорт |
Урожай- |
Зимо- |
Высота рас- |
Устойчи- |
Масса 1000 зерен, г |
Натура зерна, г/л |
Число падения, с |
Валдай (стандарт) |
68,6 |
95,0 |
152 |
4,8 |
32,5 |
709 |
112 |
Московская 12 |
72,5 |
98,4 |
147 |
5,5 |
32,1 |
708 |
187 |
От стандарта, ± |
3,9* |
3,4 |
5 |
0,7 |
-0,4 |
-1 |
75 |
* Различие достоверно на 5 % уровне значимости. |
|||||||
Эффективность отбора полносибсовых семей можно повысить, применив метод предварительного клонирования исходных растений, подлежащих взаимному переопылению. Идею использования клонов ржи в селекционных целях впервые высказал S.J. Wellensiek (13), который продемонстрировал возможность вегетативного размножения растений ржи при весеннем посеве и предложил две схемы селекции ржи с применением этого метода. В дальнейшем он модифицировал метод резервов применительно к вегетативному потомству отобранных растений, определив его как метод вегетативных половинок (14).
В 1980-е годы в Германии метод клонирования получил дальнейшее методическое развитие (15). Суть его в том, что в 1-й год проводят клонирование исходных растений (до 20 растений в клоне), затем все части клона случайно комбинируют по 2 от каждого исходного растения и проводят парные переопыления (с помощью изоляторов или стенок типа «шпанвенде»). При использовании клонов степень перекрестного опыления достигает 100 %, а от одной пары клонов можно получить достаточное число семян для проведения полевых испытаний потомств в нескольких пунктах. Новый сорт создается на базе примерно 20 лучших потомств с помощью их свободного переопыления. Метод клонирования сокращает цикл селекции на 1 год, не включает изолированное размножение полносибсовых семей, но требует много ручного труда и дополнительных финансовых затрат. Зато по эффективности его можно приравнять к линейному отбору у самоопыляющихся культур. В последние годы этот комбинированный метод полностью заменил традиционный полусибсовый отбор по причине лучшей генетической дифференциации селектируемых популяций.
Особого внимания заслуживает применение метода парных скрещиваний в первичном семеноводстве сортов. Хотя возделываемый сорт ржи в процессе семеноводства не должен претерпевать существенных изменений, полностью избежать этого нельзя из-за перекрестной природы опыления. Поэтому граница между селекцией и семеноводством сорта у озимой ржи должна быть максимально стерта. Селекционерам следует постоянно контролировать возделываемые сорта ржи: если их оставить без селекционной опеки и не проводить поддерживающей селекции, то многие ценные свойства будут постепенно трансформироваться в худшую сторону. Отбор полносибсовых семей позволяет в этой ситуации эффективно решать проблему сохранения основных достоинств сорта.
Таким образом, изложенную выше схему парных скрещиваний следует рассматривать как важный методический прием, позволяющий существенно повысить эффективность популяционной селекции озимой ржи.
Л И Т Е Р А Т У Р А
1. W o l s k i T. Metody hodowli zyta. In: Biologia Zyta. /Pod red. Cz. Tarkowskiego. Warszawa, 1983: 186-247.
2. T a r k o w s k i Cz. Genetyka, hodowla roslin, Nasiennictwo. Warszawa, 1974.
3. L a u b e W., Q u a d t F. Getreide Zuchtung. Roggen (Secale cereal L.). In: Handbuch der Pflanzenzuchtung. Bd. 2. Berlin, 1955: 35-160.
4. S e n g b u s c h R.V. Parchenzuchtung unter Ausschaltung von Inzuchtschaden. Forschungdienst, 1940, 10: 545-549.
5. N i l s s o n H. Eine Prufung der Wege und Theorien des Inzucht. Hereditas, 1937, 1-2: 236-256.
6. M u n t z i n g A. Genetical effects of duplicated fragment chromosomes in rye. Hereditas, 1943, 29: 91-112.
7. К р а с н ю к А.А. Узкородственное разведение у ржи. Тр. Саратовской оп. ст. М., 1935, вып. 1.
8. П а х о м о в а В.П. Пути повышения эффективности отбора в пределах сорта озимой ржи. Бюл. Укр. НИИ растениеводства, селекции и генетики (Харьков), 1958, 4: 10-14.
9. W o l s k i T. Studies on the inbreeding of rye. Genetica Polonica, 1971, 11(1): 1-26.
10. B u s b i c e T.H. Inbreeding in synthetic varieties. Crop Sci., 1969, 9: 601-604.
11. Г о н ч а р е н к о А.А. Определение коэффициента инбридинга в популяциях ржи, полученных при семейном отборе. С.-х. биол., 1981, ХVI(4): 552-566.
12. Г о н ч а р е н к о А.А. Реакция популяций ржи на низкий уровень инбридинга. Генетика, 1985, ХХI(2): 274-282.
13. W e l l e n s i e k S.J. Nienve methoden voor roggessele mit behulp van vegetative vermeerdering. Zaaizaad in Pootgoed, 1940, 2(7): 9-11.
14. W e l l e n s i e k S.J. Rational methods for breeding crossfertilizers. Meded Landbouwhog-geschool, 1947, 48: 227-262.
15. M i e d a n e r T. Zuchtung. In: Roggen-Getreide mit Zukunft. DLG-Verlag, 2007: 27-51.
SELECTION OF A WINTER RYE ON A METHOD OF PAIR CROSSINGS
A.A. Goncharenko
The scheme of selection process of a winter rye with use of a method of pair crossings is offered. Selection advantages of the new scheme are analyzed, a level inbreeding in the populations of the rye received at association separately multiplied halfsib and fullsib of posterities is dismissed. It is shown, that arising inbreeding it is possible to lower to a zero level by free transpollination corresponding quantity of families. Minimally admissible number of parental components at pair crossings should make nearly 40.
Key words: winter rye, method of reserws, pair crossing, full sib family, coefficient of inbreeding.
ГНУ НИИ сельского хозяйства центральных |
Поступила в редакцию |




